Seaweed-Derived Sulfated Polysaccharides; The New Age Chemopreventives: A Comprehensive Review
Sep 22, 2023
Simple Summary: Sulfated polysaccharides are powerful chemotherapeutic or chemopreventive agents that have anti-cancer properties by increasing immunity and driving apoptosis in several cancer cell lines. Sulfated polysaccharides have significant antioxidant and immunomodulatory potentials, which contribute to their disease-preventive effectiveness with low cytotoxicity and good efficacy therapeutic outcomes in cancer via dynamic apoptosis modulation. Furthermore, it can be used as a dietary supplement or as an adjuvant treatment for cancer.

Benefits of cistanche tubulosa-Antitumor
Abstract: Seaweed-derived bioactive compounds are regularly employed to treat human diseases. Sulfated polysaccharides are potent chemotherapeutic or chemopreventive medications since it has been discovered. They have exhibited anti-cancer properties by enhancing immunity and driving apoptosis. Through dynamic modulation of critical intracellular signaling pathways, such as control of ROS generation and preservation of essential cell survival and death processes, sulfated polysaccharides’ antioxidant and immunomodulatory potentials contribute to their disease-preventive effectiveness. Sulfated polysaccharides provide low cytotoxicity and good efficacy therapeutic outcomes via dynamic modulation of apoptosis in cancer. Understanding how sulfated polysaccharides affect human cancer cells and their molecular involvement in cell death pathways will showcase a new way of chemoprevention. In this review, the significance of apoptosis and autophagy-modulating sulfated polysaccharides has been emphasized, as well as the future direction of enhanced nanoformulation for greater clinical efficacy. Moreover, this review focuses on the recent findings about the possible mechanisms of chemotherapeutic use of sulfated polysaccharides, their potential as anti-cancer drugs, and proposed mechanisms of action to drive apoptosis in diverse malignancies. Because of their unique physicochemical and biological properties, sulfated polysaccharides are ideal for their bioactive ingredients, which can improve function and application in disease. However, there is a gap in the literature regarding the physicochemical properties and functionalities of sulfated polysaccharides and the use of sulfated polysaccharide-based delivery systems in functional cancer. Furthermore, the preclinical and clinical trials will reveal the drug’s efficacy in cancer.
Keywords: apoptosis; cancer; chemoprevention; sulfated polysaccharides
1. Introduction
The current global population explosion and altered dietary and lifestyle practices are considered critical factors for disease occurrence. Numerous infection-driven diseases, along with Alzheimer’s, Parkinson’s, diabetes, cancer, and other neurological disorders, pose a severe risk to the human lifespan [1]. Cancer, a collection of numerous pathological problems brought on by unchecked cell growth, has detrimental effects on individual health care [2]. According to estimates from 2019, there are more than 200 deadly cancer kinds that cause more than 9.6 million deaths annually worldwide [3]. The leading causes of death are skin, stomach cancer, breast, lung, prostate, and colorectal cancer [4]. Instances of mortality in low- and middle-income nations are documented in about 70% of cases [5]. Based on epidemiological studies, the World Health Organization (WHO) predicted 9.6 million cancer-related deaths and 18 million new cases in 2018 [6]. Several homeostatic systems are disrupted by the uncontrolled proliferation of cancer cells leading to their invasiveness and metastasis owing to genetic changes [7]. Cancer treatment methods include surgery, chemotherapy, radiation therapy, and immunotherapy; chemotherapy is the most frequently used [7–9]. Chemotherapy is a common and efficient cancer treatment that damages several important organs by causing cytotoxicity in both cancerous and non-cancerous cells [10]. Drug tolerance is the main concern in cancer treatment in order to eliminate side effects and severe reactions [11,12]. Therefore, it is crucial to develop and look for anti-cancer drugs with fewer side effects and higher tolerance. Chemotherapy sometimes creates an unfavorable setting and irreversible organ damage surrounding the target site. Additionally, cellular lenience to the medications poses additional therapeutic difficulties. Therefore, it is desired to find fresh therapeutic agents with low side effects to abide all the adverse conditions [13]. The naturally occurring bioactive compounds used as drugs have a variety of therapeutic applications [14–18]. Additionally, most medicinal drugs are either natural compounds or synthetic equivalents [19]. Due to their varied chemical makeup and bioavailability, marine natural products (MNPs) have recently been investigated for their potential as therapeutic candidates [20–25]. Algal biodiversity is abundant in marine and freshwater settings and contributes to the main bioactive metabolites [24,26–36]. Seaweeds are found in both freshwater and saltwater, and they play a significant part in preserving the ecology and biodiversity of marine ecosystems [37,38]. Anticoagulant, anticancer, antidiabetic, antiviral, immunomodulatory, antiangiogenic, anti-inflammatory, antiadhesive, and anti-neurodegenerative properties of sulfated polysaccharide use as potential therapeutic agents [39–44]. Fucoidan, porphyrin, carrageenan, and ulvan are sulfated polysaccharides often extracted from brown, red, and green algae and contain sulfate groups that have the possibility as therapeutic diligences against many malignancies [45–47]. Sulfated polysaccharides are expected to be used as chemotherapeutic pharmacological agents in clinical practice due to their enormous structural variety and robust antioxidant capacity [48]. Additionally, its high absorption, cheap maintenance costs, improved production, and use as food supplements make it a more sought-after chemotherapy drug [49]. Previous reports have discussed the anticancer properties of algal-derived sulfated polysaccharides [50–52]. However, the exact mechanism is not fully understood or discussed. Moreover, the context-specific drug targets, cancer subtypes and tumor microenvironment are not discussed [52–54]. The present form of the manuscript discusses the mechanistic involvement of these algal-derived sulfated polysaccharides in the induction of cell death pathways. The multi-target specific one-drug therapy has also been discussed keeping the tumor heterogeneity in mind. The systematic analysis of the origin and mechanistic overview of the sulfated polysaccharide regulatory pathways used in cancer prevention are the main topics of this review. To comprehend therapeutic intervention in the context of cancer prevention, we have also concentrated on the chemical complexity and sources of sulfated polysaccharides. Future clinical and nano drug delivery uses are suggested by this review, which also takes into account the potential function of sulfated polysaccharides in cancer prevention.
")
Chinese herb cistanche plant-Antitumor
2. Intricate Role of Apoptosis in Cancer Treatment: The Programmed Cell Death
Understanding the pathogenesis of diseases brought on by apoptosis dysfunction requires an understanding of the mechanisms of apoptosis. The creation of medications that specifically target apoptotic genes or pathways may benefit from this. Because they function as both initiators and executors, caspases are essential parts of the apoptosis mechanism. There are three distinct pathways by which caspases can be activated. Intrinsic (or mitochondrial) and extrinsic are the two apoptosis initiation pathways that are most frequently discussed (or death receptor) (Figure 1). Both pathways ultimately lead to the execution stage of apoptosis, which is a common pathway. The intrinsic endoplasmic reticulum pathway is a third, less well-known initiation pathway [55].

Figure 1. Role of apoptosis in cancer treatment
Numerous caspases are activated during the apoptosis execution stage. Caspase 9 mediates the intrinsic pathway, while Caspase 8 mediates the extrinsic pathway. Both intrinsic and extrinsic pathways converge on caspase 3 to complete apoptosis. Nuclear apoptosis is caused by the degradation of the caspase-activated deoxyribonuclease inhibitor by caspase 3 [56]. Additionally, protein kinases, cytoskeletal proteins, DNA repair proteins, and endonuclease inhibitory subunits are cleaved by downstream caspases. Additionally, they influence the cytoskeleton, cell cycle, and signaling pathways, all of which help to shape the specific morphological changes that take place during apoptosis [56]. Apoptosis is an energy-dependent programmed cell death characterized by membrane blebbing, shrinking cytoplasmic chromatin condensation, and nuclear disintegration. Apoptosis is the process by which cells die without causing inflammation [57,58]. Additionally, it can be started by mitochondrial-mediated mechanisms or surface death receptors (DR; extrinsic apoptosis) (intrinsic apoptosis) [59–62]. Both pathways cause executive caspases to be activated, which cleave molecules related to the structural and regulatory molecules of the network of apoptotic cells [46,63,64]. After pathogenic stressors, apoptosis is a cell death mechanism that aids in maintaining cellular homeostasis [65]. Malignant cells typically go through a series of genetic mutations to survive pathogenic stimuli. Apoptosis resistance or decreased apoptosis promotes carcinogenesis [66–68]. Cancer cells frequently avoid apoptosis by rebalancing the pro- and anti-apoptotic protein balance. Cancer cells can also avoid apoptosis if their caspase activity is low and their DR signaling is compromised [62,69,70]. Bcl-2 family proteins, an inhibitor of apoptosis proteins (IAPs), p53, executioner caspases, and DRs are frequently affected by cancer cells. These molecular genes and their related pathways are critical in cancer therapies because they cause apoptotic cell death [71–75]. Apoptosis’s typical role in cancer treatment is depicted (Figure 1).
3. Seaweeds: The Chief Contributor of Sulfated Polysaccharides
Due to their enormous biodiversity and use as food and traditional medicine worldwide, seaweeds are thought to be a good source of bioactive chemicals [76–78]. The therapeutic effects of a number of seaweed-derived bioactive chemicals, their unprocessed extracts, and partially purified polysaccharides on a range of human diseases have been investigated [54,79,80]. Their antioxidant qualities aid the ability of the phytoproducts made from seaweed to resist disease. Brown seaweeds have various physical and functionally distinct polysaccharides, including alginic acids, and fucoidans [81,82]. In biotechnology, medicine, and food preparation, sulfated polysaccharides are frequently used [83,84]. Polyphenols, free amino acids, iodine-containing substances, vitamins, and lipids isolated from seaweeds are examples of low molecular metabolites used in food processing and medicine [81,85]. The sulfated polysaccharides are physiologically active, highly branched, different from monosaccharide composition, and have a higher molecular weight. Long chains of linked sugar molecules make up the fucoidan, which is decorated with sulfate groups [86]. Sulfated polysaccharides’ ability to fight off many malignancies is mainly attributed to their antioxidant capability [87–89]. It is well known that the sulfated polysaccharides derived from seaweeds are effective anticancer drugs. Seaweed contains a variety of sulfated polysaccharides. According to their chemical makeup, polysaccharides are categorized as Galatians, and sulfated xylans, sulfuric acid polysaccharides (generally found in green algae). Brown algae also contain fucoidan [90]. Red algae frequently contain agar, carrageenans, xylans, and floridean. Several algal sulfated polysaccharides could be used as therapeutic candidates to address a variety of human health inequalities [91]. Sulfated galactans known as carrageenans are frequently used in the food and drug industries. soluble fibers, like fucans, are found in brown seaweeds. On the other hand, red seaweeds are abundant in soluble fibers like xylans, Floridian starch, and sulfated galactans (agars and carrageenans) [92]. Green algae also contain xylose, galactose, uronic acids, arabinose, and rhamnose, as well as mannans, xylans, starch, and polysaccharides with ionic sulfate groups. There are many types of soluble and insoluble fibers in polysaccharides [93,94]. Compared to their dry weight, seaweeds provide a more significant percentage of dietary fibers (between 25% and 75%) than those found in fruits and vegetables [95]. Consuming dietary fiber from algae has many positive health effects since it acts as an antitumor, anticancer, anticoagulant, and antiviral agent. In brown macroalgae, sulfated polysaccharides are extensively distributed in the cell walls [80]. Among other biological actions, sulfated polysaccharides act as an antioxidant, anti-inflammatory, anticoagulant, anticancer, antiviral, antidiabetic, and antithrombotic agent. They also alter the human immune system [7]. Additionally, fucoidan, which is rich in brown seaweeds and is the second largest source of sulfated polysaccharide, promotes intestine metabolism in human health [94].
The Structural Complexity of Seaweed-Derived Sulfated Polysaccharides
Research is increasingly focusing on polysaccharides, which are found in seaweeds and have anticancer, antioxidant, anti-coagulant, and anti-inflammatory properties [7,96]. Polysaccharides are large molecules classified by a monomeric unit as homopolysaccharides, homoglycans, heteropolysaccharides, or heteroglycans. Polysaccharides are also classified according to their seaweed origin as brown, red, green, or blue. Fucoidan (a sulfated polysaccharide), is the main component of brown seaweed. Agars, xylans, carrageenans, Floridian starch (glucan that resembles amylopectin), water-soluble sulfated galactan, and porphyrin are some products made from red algae. Green seaweeds contain sulfated galactans, xylans, and polysaccharides. Seaweed contains a variety of polysaccharides, with some genera—including Ascophyllum, Porphyra, and Palmaria—containing up to 76% polysaccharide by dry weight [97].
4. Disease Preventive Activity of Sulfated Polysaccharides: The Magic Bullets
Numerous studies have demonstrated that the biological activity of polysaccharides is influenced by their molecular weight, conformational state, chemical composition, and glycosidic connections [98]. Understanding the relationship between molecular weight and essential properties, such as polysaccharide viscosity, conformation, water solubility, and others, is important in cancer [99,100]. Porphyrins with a lower molecular weight (LMW) have more potent antioxidant properties [101,102]. Since porphyran has a lower mean molecular mass, it has a more significant ROS-scavenging activity [103]. Additionally, the byproduct of porphyrin acid hydrolysis, oligo-porphyrin, has the potential to both prevent and treat a number of cancers. A higher irradiation exposure dose and porphyrin with a lower molecular weight were required because gamma radiation damaged the anti-cancer response of porphyrin derived from P. yezoensis [45]. Contrary to earlier research that claimed lower molecular weight porphyrin has more potent anti-cancer activity, porphyrin inhibited cancer cell lines HeLa and Hep3B more potently than the degraded products. The composition of the monosaccharide or sulfate did not change significantly [100]. Therefore, future research should focus on how the molecular weight of porphyrin and its anti-cancer activity are related. In Asian nations like Japan, China, Thailand, and South Korea, edible seaweeds are valued as a wholesome food source. The fight against cancer necessitates the use of polyphenols, terpenes, phycobiliproteins, carotenoids, phlorotannins, pigments, and polysaccharides [104]. Antioxidants found in seaweed’s anti-cancer properties help stop the spread of cancer. As cancer progresses, antioxidants are crucial because they inhibit tumor growth without causing cytotoxicity [105]. For instance, a mouse model of sarcoma 180 was successfully treated with an immune-stimulating sulfated polysaccharide from Champia feldmannii without cytotoxicity [106]. The polysaccharides from the Gracilaria lemaneiformis induced splenocyte proliferation, macrophage phagocytosis, and tumor inhibition. Mice with H22 hepatoma cell transplants had higher levels of IL-2 and CD8+ T lymphocytes in their blood [107]. A sulfated polysaccharide from C. feldmannii showed anti-cancer efficacy in Swiss mice in vitro and in vivo. Increasing the production of OVA-specific antibodies improves immunity [106]. Fucoidans’ anti-cancer properties have been confirmed in a diversity of cancers, including stomach, breast, lung, and liver cancers [7]. Fucoidan has received more attention than porphyrin and other sulfated polysaccharides. Sulfated polysaccharides from green, brown, and red algae have sparked a lot of interest in this context due to their anticancer properties. The physicochemical characteristics of the different sulfated polysaccharides and their wide range of therapeutic potential (Table 1) will be leading this research. Some physicochemical properties of sulfated polysaccharides have been reported, including ionic solubility, crosslinking, biocompatibility, nontoxicity, rheological properties, and biodegradability [108–110]. These properties are important characteristics of sulfated polysaccharides that have sparked a lot of interest in their application. Fucoidan’s primary properties are ionic crosslinking and solubility [109]. The water-soluble sulfated polysaccharides facilitate the development of fucoidan and other positively charged molecule-based delivery systems. Fucoidan’s negatively charged sulfate groups, for example, could be communal with chitosan’s ammonium groups to form nanoparticles, hydrogels, and comestible films for nutraceutical delivery [111,112]. Nontoxicity is an important property in addition to ionic crosslinking and solubility. Except as previously stated, biodegradability and biocompatibility are critical factors in facilitating the use of sulfated polysaccharides in therapeutic and drug delivery systems. Researchers have recently become interested in sulfated polysaccharides because of their excellent biocompatibility and biodegradability. The biodegradability of sulfated polysaccharides can increase the bioavailability and delivery effectiveness of bioactive ingredients. Depolymerization and purification can generally increase the biodegradability of sulfated polysaccharides by lowering their molecular weight, but this method is too expensive to be widely used [113].
Table 1. The physicochemical characteristics of sulfated polysaccharides and their therapeutic potential with other functions.

Apoptosis Modulatory Seaweeds Derived Sulfated Polysaccharides
As anticancer anti-angiogenic, and anti-inflammatory drugs, sulfated polysaccharides have a variety of biological effects [43,114]. Sulfated polysaccharides isolated from different marine habitats have been extensively studied and found to be effective anticancer mediators against various cancer cell lines by modulating numerous cell survival pathways and inducing apoptosis [43,46]. Therefore, Sulfated polysaccharides secluded from various green, brown, and red seaweeds from various marine habitats have been extensively studied for their ability to dynamically regulate cell death pathways. These polysaccharides are effective anticancer agents because they modulate numerous cell survival pathways and induce apoptosis. Sulfated polysaccharides are widely used in cancer therapies as well as precision medicine to develop next-generation drugs. Green seaweeds: Green algae, also known as Chlorophyta, are an influential group of marine algae that are a source of polysaccharides [115]. However, green algae cell wall polysaccharides have received less attention than red (agarans and carrageenan) and brown algal polysaccharides (fucoidan) [96]. Nonetheless, the study of sulfated polysac charides derived from green seaweeds has piqued the scientific community’s interest in recent years, primarily because of their structural diversity, and biological, and physicochemical properties [116]. Furthermore, due to their variety of glycosidic linkages that result in branched structures and attached sulfate groups with various special distributions, sulfated polysaccharides are the most bioactive and promising candidates [117]. Ulvan are sulfated polysaccharides that are water-soluble and derived from the cell walls of green algae. They are present in plants belonging to the genera Ulva, Enteromorpha, Monostroma, Caulerpa, and others. They consist of repeating disaccharide moieties like sulfated rhamnose and uronic acid (glucuronic or iduronic). Glycosaminoglycans, which are present in the extracellular matrix of animal connective tissues, have a structure that is comparable to that of Ivan disaccharide moieties. Some even also have xylose residues visible (Figure 2) [118]. Highly pyruvate 1,3-D-galactan sulfate from the Codium expense and a polysaccharide similar to it from Codium is thmocladium are two other types of polysaccharides found in green algae [119,120]. Sulfated β-D-mannans have also been discovered, such as those isolated from Codium vermilara [121]. The molecular structure of van sulfated polysaccharide is displayed in Figure 2.

Figure 2. The Molecular structure of Ivan is drawn in ChemDraw 12.0 Ultra.
Sulfated polysaccharides isolated from various tropical green algae have recently been found to have antioxidant and antiproliferative properties. After 72 h of incubation, HeLa cell proliferation was reduced by 36.3% to 58.4% by the polysaccharide isolated from Caulerpa prolifera [122]. Two polysaccharide fractions from the Caulerpa racemosa, a green alga, showed antitumor activity at a dose of 100 mg/kg/day, with inhibition rates of H22 tumor transplanted in mice of 59.5–83.8% (48 h) and 53.9% (14 days), respectively [123]. Through in vivo and in vitro experiments, water-soluble sulfated polysaccharide fractions of Enteromorpha prolifera were found to stimulate immunity. These polysaccharides significantly increased ConA-induced splenocyte proliferation and cytokine production through elevated m-RNA expression [124]. Ulvan from Ulva rigida stimulated the secretion and activity of murine macrophages, increased COX-2, and NOS-2 expression, and more than doubled the expression of some cytokines [125]. Ulvans from Ulva pertusa stimulated nitric oxide and cytokine production while causing little cytotoxicity against tumor cells [126]. Several studies on the antioxidant activity of Ivan in experimental D-galactosamine-induced hepatitis in rats have been published [127,128]. Polysaccharides derived from green algae have potent immunomodulatory and antioxidant properties, implying that they could be used to prevent cancer. Ulvan’s anticancer activity has recently been discovered in U. australis, U. lactuca, U. ohnoi, and U. rigita [129]. Several studies have investigated ulvan in toxicity and cell viability to test its anticancer activity, specifically for anti-breast cancer, anti-colon, and anti-cervical cancer activity [129–132]. Ulvan contains sulfated polysaccharides, which inhibit the proliferation of hepatocellular carcinoma and induce apoptosis. By lowering oxidative stress, sulfated polysaccharides protect the liver from DNEA-induced damage [133]. Additionally, they enhance apoptosis, reduce oxidative stress and inflammation, and strengthen the antioxidant defense system in DMBA-treated mice [130]. Ulvan was less toxic to A459 and LS174 cells (IC50 > 200 mg/mL), but it was more effective against Fem-x and K562 cells (IC50 74.73 and 82.24 mg/mL, respectively) when it came to preventing moderate cytotoxicity [134]. With IC50 values ranging from 21 to 99 µg/mL, even reduced tumor growth in MCF-7 and HCT-116 cells [132] and strong ligand bonds appear to connect this to sulfated polysaccharides [135]. Ulvan inhibited the growth of hepatocellular carcinoma (IC50 29.67 ± 2.87 µg/mL), human breast cancer (IC50 25.09 ± 1.36 µg/mL), and cervical cancer (IC50 36.33 ± 3.84 µg/mL) [131]. However, Caco-2 cell proliferation or differentiation can be inhibited by low molecular weight polysaccharides (5000 Da), usually oligosaccharides [129]. Sulfated polysaccharides have an antiproliferative effect, but it depends on the cell type. Sulfated polysaccharide TPs (precipitated in alcohol) extracted from the green alga Codium Bernabei exhibited low cytotoxicity on HCT-116 and MCF-7 cell lines in comparison to APs (precipitated in acid media). On the other hand, the HL-60 cell lines showed little cytotoxicity when exposed to the APs [51]. Due to its strong antioxidant activity, Enteromorpha spp. the extract has antiproliferative effects on cancer cell lines like Fem-x, A549, LS174, and K562 [136]. Additionally, a different solvent extract of Enteromorpha compress extract induces anticancer activity via apoptosis in oral cancer cell lines Cal33 and FaDu [38]. Brown seaweeds: Brown seaweeds are the most promising sources of sulfated polysaccharides and displayed the most promising anticancer activity against various cancer cell lines. The typical sulfated polysaccharide structure derived from brown seaweeds is displayed (Figure 3). Lewis lung cancer cells (LCC) and melanoma B16 cells were discovered to be sensitive to the fucoidan isolated from Sargassum sp. [137]. It reduced cell proliferation and dose-dependently promoted apoptosis, as shown by morphological alterations. The fucoidans from S. hemiphyllum inhibited the growth of breast cancer by upregulation of miR-29c and downregulation of miR-17-5p. Furthermore, it was clear that after fucoidan administration, EMT progression was slowed by amplified E-cadherin and reduced N-cadherin expression. Furthermore, activation of the pathway of phosphoinositide 3- kinase/Akt has promoted apoptosis in breast cancer cells [138]. Fucoidan from L. gurjanovae demonstrated an anti-neoplastic effect in rat epidermal JB6 Cl41 cells by delaying EGFR phosphorylation. It controlled EGF-induced c-jun signaling and inhibited the action of activator protein-1 (AP-1) [139].

figure 3. The Molecular structure of different types of sulfated polysaccharides such as fucoidan, with potential therapeutic effects, are drawn in ChemDraw 12.0 Ultra.
Fucoidan derived from F. vesiculosus inhibited cell proliferation and arrested the cell cycle in ovarian cancer (ES2 and OV90) cells. It also produced ROS, which regulated intrinsic apoptosis. By suppressing the PI3K and MAPK signaling pathways, ER stress also promoted apoptosis. It also demonstrated anticancer effects on human mucoepidermoid carcinoma by modifying the p-38 MAPK, ERK1/2, and JNK pathways (MC3) [140]. Further, it reduced the amount of calcium in the cytosol and mitochondria to support apoptotic cell death. Similar extraction techniques for fucoidan produced from F. vesiculosus showed in vivo anticancer efficacy in the zebrafish xenograft and fli1 Tg model [141]. In HepG2 and HeLa G-63 cells, fucoidan from Fucus vesiculosus demonstrated potent anticancer activity. Fucoidan was discovered to be more effective in human liver cancer cells (HepG2) [142]. Fucoidan from Fucus vesiculosus increased MMP, which induced caspase-3-dependent apoptosis in human Burkitt’s lymphoma (HS-Sultan) cells. Furthermore, reports of caspase-independent apoptotic cell death in HS-Sultan cells were seen after fucoidan administration. Additionally, fucoidan prevented the ERK and GSK pathways from being phosphorylated, both of which were necessary for the activation of apoptosis [143]. Its low IC50 (34 µg/mL) activated pro-caspase-3, pro-caspase-9, and caspase-3/7 while downregulating Bcl-2 in HCT-15 cells [144]. The ability of anti-apoptotic proteins like Bcl-xl, Bcl-2, and Mcl-1 to cause apoptosis in MDA-MB231 cells was inhibited by fucoidan at IC50 (820 µg/mL) [46]. Fucoidan (IC50; 20 µg/mL) therapy led to a similar fluctuation in the expression of Bad, Bcl-2, Bim, Bcl-xl, and Bik in colon cancer cell lines [144]. Fucoidan derived from C. okamuranus was combined with Con A, and it promoted intrinsic apoptosis by caspase-3/7 induction in HL60 cells [145]. In addition, glutathione depletion and NO production were significant mediators of apoptosis in human leukemia cells, as were the activation of MEKK1, ERK1/2, MEK1, and JNK [146]. Fucoidan from C. novaecaledoniae was extracted and used to induce intrinsic apoptosis in HeLa, MCF-7, MDA- MB-231, and HT1080 cells. This intrinsic apoptosis was accompanied by MMP, DNA fragmentation, nuclear condensation, and phosphatidylserine externalization [147]. C. okamuranus fucoidan induced caspase-dependent apoptosis in U937 cells by inducing the caspase-3 and -7 pathways [148]. Additionally, it enhanced mouse cell-mediated immunity, phagocytes, and immune cell proliferation in the in vivo model [149].
")
cistanche supplement benefits-increase immunity
Additionally, in normal stomach (Hs 677. St) cells, fucoidan isolated from C. okamuranus reduced cellular damage brought on by 5-fluorouracil (5- FU) [150]. In this setting, significant anti-proliferative activity in MCF-7 cells was observed, with no cytotoxicity to human mammary epithelial cells. There was an increase in caspase-7, caspase-8, and caspase-9 activity, internucleosomal DNA fragmentation, and chromatin condensation in both cell lines [151]. Fucoidan therapy has also been reported for caspase-independent cell death in MCF-7 [152]. Hydrolyses increase the luminal fucoidan content, which is a potent chemopreventive mediator of colon cancer because they do not digest these fucoidans [153]. Fucoidan (0–20 µg/mL) therapy promoted mitochondrial death in HT-29 and HCT116 cells via caspase-3 regulation. Extrinsic apoptosis in HT-29 cells has also been reported recently [144]. The anticancer properties of C. okamuranus low molecular weight fucoidan (LMWF; 6.5–40 kDa), high molecular weight fucoidan (HMWF; 300–330 kDa), and intermediate molecular weight fucoidan (IMWF; 110–138 kDa) were demonstrated in a colon carcinoma tumor-bearing rat model [154]. Fucoidan (MW 5100 kDa) from U. pinnatifida induced apoptosis in human prostate cancer (PC-3) cells via induction of ERK1/2 MAPK, inhibition of p38 MAPK, and PI3K/Akt pathway. Furthermore, the downregulation of the Wnt/-catenin pathway aided the progress of apoptosis [155]. Fucoidan also amplified the p21Cip1/Waf pathways in PC-3 cells. Furthermore, it reduced E2F-1 cell cycle-related proteins while increasing Wnt/-catenin pathways. GSK-3 activation reduced the expression of c-MYC, and cyclin D1, which aided anti-proliferative activity [156]. These fucoidans were found to have anticancer activity in HeLa, A549, and HepG2 cells by altering the previously mentioned critical cellular signaling pathways [157]. Fucoidan from F. vesiculosus induced apoptosis in cancer cell lines including NB4, THP-1, and HL-60. Fucoidan administration activated caspases-3, -8, and -9, cleaved Bid, and altered MMP in HL-60 cells. The initiation of apoptosis had a comparable effect in U937 cells. Moreover, in U937 cells, fucoidan therapy increased MMP (mitochondrial membrane potential) and cytosolic cytochrome C release, as well as the Bax/Bcl-2 ratio. Caspase inhibitors, on the other hand, delayed the onset of apoptosis, demonstrating that fucoidan-regulated caspase activity was accountable for apoptosis induction. Furthermore, treatment with SB203580, a specific p38 MAPK inhibitor, was accountable for apoptosis discount, demonstrating the importance of MAPK in activating apoptosis [158]. Fucoidan therapy inhibited the G1 cell cycle in EJ cells by affecting cyclin D1, cyclin E, and Cdks (cyclin-dependent kinases). Furthermore, it inhibits Rb phosphorylation, which results in cellular aging [159]. Fucoidan from F. vesiculosus inhibited the growth of MCF-7 cells by stopping the cell cycle at the G1 phase and lowering CDK-4 and cyclin D1 levels. Furthermore, by cleaving PARP and Bid, decreasing Bcl-2, and increasing Bax, it induced ROS-dependent apoptosis. MCF-7 cells exhibited the onset of intrinsic apoptosis via regulation of caspase-7, -8, and -9 and cytosolic cytochrome C release [160,161]. Furthermore, fucoidan from F. vesiculosus therapy reduced cell migration and invasion as well as EMT in MCF-7 cells by downregulating MMP-9 and overexpressing E-cadherin [162]. Fucoidans derived from F. vesiculosus inhibited growth in MDA-MB-231 and 4T1xenograft female Balb/c mouse cells, resulting in less metastatic lung nodule development. The effective setback of TGFR induced EMT was achieved mechanistically by downregulating TGFRII and TGFRI. The cases mentioned above have all been associated with the upregulation of epithelial markers and their phosphorylation of Smad2/3 Smad4 expression, phosphorylation of Smad2/3 Smad4 expression, and downstream signaling molecules [163]. Furthermore, caspase-3 activation, cytosolic cytochrome C release, downregulation of Bcl-2, and increased Bax expression induced apoptosis. In addition, the regulation of VEGF, Survivin, and ERKs expression aided in the commencement of apoptosis [164]. In the MDS/AML and SKM1 cell line, treatment with marketed fucoidan (100 µg/mL for 48 h) caused cell cycle arrest (G1 phase) and Fas instigation to induce extrinsic apoptosis via caspase 8 and 9 modulations. Furthermore, it influenced PI3K/Akt pathway in a ROS-dependent manner, thereby promoting apoptosis [165]. It altered p-Akt, p-PI3K, p-P38, and p-ERK, to modulate MAPK and PI3K/Akt signaling pathways in DU-145 cells (prostate cancer). Furthermore, it increased Bax expression while decreasing Bcl-2, PARP cleavage, and caspase-9 expression in a concentration-dependent manner [166]. Fucoidan administration induced apoptosis in osteosarcoma (MG-63) cells (evidenced by cellular blabbing, nuclear disintegration, and chromatin condensation) [167]. Treatment with marketed synthetic fucoidan increased ROS-regulated apoptosis in human bladder cancer (5637) cells by activating mitochondrial membrane potential (MMP), increasing the Bax/Bcl-2 ratio, and increasing cytosolic cytochrome C release. Furthermore, inhibition of PI3K/Akt signaling and anti-telomerase activities promoted apoptotic cell death in 5637 human bladder cancer cells via downregulating telomerase Activity [168]. Furthermore, AKT signaling activation was claimed to be critical in inhibiting proliferation and suppressing bladder cancer cells’ ability to migrate and invade [169]. Fucoidan inhibited the cell cycle in 5637 and T-24 cells (human bladder carcinoma) by altering the expression of p21/WAF1, cyclins, and CDK. Furthermore, MMP-9 inhibition via AP-1 and NF-kB reduced bladder cancer cell proliferation [169]. Sulfated polysaccharides from brown algae as potent anticancer agents are displayed in Table 2.
Table 2. Sulfated polysaccharides from brown algae as potent anticancer agents. ↑: upregulation, ↓: Downregulation, ⊥: Inhibition



Red seaweeds: Porphyran is a polymer found in Porphyra sp., a red seaweed. The porphyran is a galactose that has been heavily replaced by L-galactose 6-O-sulfation and 6-O-methylation [100]. The typical repetitive structure of porphyran is displayed (Figure 4). Porphyran is extracted from red seaweeds using hot water extraction, ultrasonic treatment, and radical degradation. Human studies have demonstrated the anticancer, hypolipidemic, and anti-inflammatory properties of porphyran [170]. When consumed orally, porphyran shields the livers of ICR mice from the effects of a high-fat diet, suggesting that it might be used as a dietary hypolipidemic component [171].

Figure 4. The Molecular structure of different types of porphyran with potential therapeutic effects are drawn in Chemdraw 12.0 Ultra.

Figure 5. The Molecular structure of different types of carrageenan with potential therapeutic effects is drawn in ChemDraw 12.0 Ultra. The Polymeric structures of the different molecular structures of carrageenan as Υ-carrageenan (a); β-carrageenan (b); δ-carrageenan (c); α-carrageenan (d); µ-carrageenan (e); κ-carrageenan (f); ν-carrageenan (g); ι-carrageenan (h); λ-carrageenan (i); and θ-carrageenan (j).
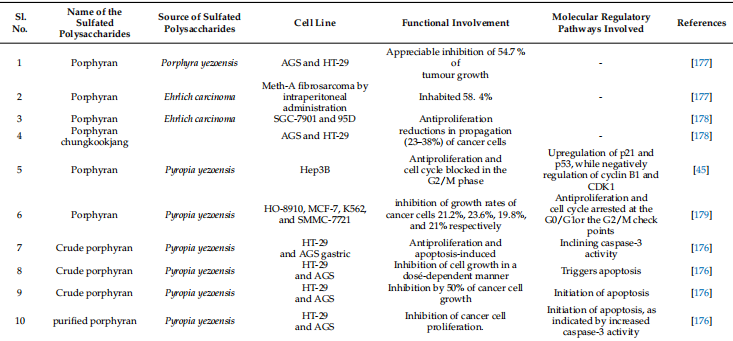
Cancer is known to be accelerated by free radicals and ROS (reactive oxygen species). Synthetic chemopreventive drugs usually generate undesirable side effects in the tumor environment due to their low selectivity and extensive biodistribution [173]. Porphyrin is a potent chemopreventive agent due to its influence on cellular proliferation, the cell cycle, and the induction of apoptosis [174]. The red alga Porphyra yezoensis can induce apoptotic cell death in cancer cell lines in vitro while causing no cytotoxicity to normal cells. Generally speaking, porphyrin is not toxic to healthy cells, but it is toxic to cancer cells, leading to dose-dependent cell death [175]. Additionally, it has been demonstrated that porphyrin inhibits overall cell growth while inducing apoptosis in AGS human stomach cancer cells [175]. In AGS cells, the insulin-like growth factor-I receptor/Akt pathway increases PARP cleavage and caspase-3 activation, which encourages cell death [175]. Numerous studies have demonstrated the antitumor and anticancer properties of porphyrin and its oligosaccharides. Porphyrin can encourage the cleavage of poly (ADP-ribose) polymerase and the activation of caspase 3 in gastric cancer cells. By reducing the expression levels in AGS cells (gastric cancer), porphyrin may slow the growth of cancer cells. This would then prevent IGF-IR phosphorylation and activate caspase 3 [175]. Crude and purified porphyrin have antiproliferative activity in HT-29 and AGS cells in vitro. Apoptosis is induced by the crude porphyrin polysaccharide component, as shown by an increase in caspase-3 activation [176]. Porphyrin inhibits HT-29 cell proliferation by activating caspase-3 [176]. Porphyrin has been shown to be effective against Ehrlich cells (EAC) carcinoma and Meth-A fibrosarcoma in mouse tumor models [177].

Desert living cistanche tubulosa
Click here to view Cistanche products
【Ask for more】 Email:cindy.xue@wecistanche.com / Whats App: 0086 18599088692 / Wechat: 18599088692
Natural porphyrin was found to have no effect on MDA-MB-231, whereas two breakdown products had an impact when porphyrin and two OPs (Oligo-porphyrin) created by gamma irradiation were tested for anticancer activity. By preventing the cell cycle from entering into the G2/M phase, OPs have the capacity to reduce the growth of cells [45]. As a result, porphyran’s MW has displayed a significant impact on its anticancer efficacy. Low-MW of OPs are particularly effective against cancer; however, macromolecular porphyrin has no antitumor activity. Furthermore, the anticancer activity of porphyrin was discovered, with porphyrin mainly acting as an anticancer drug by hindering cell growth and tempting apoptosis [180]. Carrageenans have been shown in numerous studies to have antiproliferative activity in cancer cell lines in vitro and tumor growth inhibitory effectiveness in mice [181–183]. They also have an antimetastatic effect by preventing cancer cells from connecting with the basement membrane and limiting tumor cell propagation and adhesion to different substrates; however, the precise mechanisms of action are yet unknown. Carrageenans from Kappaphycus alvarezii were found to prevent the growth of cancer cells from the liver, colon, breast, and osteosarcoma [184]. Yamamoto et al. (1986) discovered that taking various seaweeds orally significantly reduced the occurrence of carcinogenesis in vivo [185]. Hagiwara et al. (2001) [186] examined carrageenan’s effects on colonic carcinogenesis in male rats. Treatment had no effect on clinical symptoms or body weight. According to histological research, carrageenan has no colorectal carcinogenesis encouraging activity at the maximum dietary intake of 5.0% in the existing experimental settings [186]. Carrageenan has been shown in several studies to have specific cytotoxic effects on cancer cells. In such studies, doses of 250–2500 µg/mL of both k-carrageenan and λ-carrageenan inhibited human cervical cancer cells by stopping the cell cycle at specific stages and delaying its completion [47]. k-carrageenan delayed the cell cycle’s (G2/M stage), whereas λ-carrageenan delayed both the G1 and G2/M stages. However, k-selenocarrageenan (selenocarrageenan containing selenium) inhibits cell propagation in a human hepatoma cell. The cell cycle is terminated during the S phase of the cell cycle [187]. In vivo and in vitro studies, however, exposed that native carrageenan had no discernible anti-proliferation effect in the human osteosarcoma cell line. Because of a reduction in the Wnt/-catenin signaling pathway, degraded carrageenan-induced apoptosis inhibited tumor growth, and stopped the G1 phase of the cell cycle, all of which increased the existing rates of tumor-bearing mice [188]. Angiogenesis is a critical step in the progression of cancer. As a result, anti-angiogenic activity in cancer treatment is being extensively researched. Carrageenans are angiogenesis inhibitors due to their higher anti-angiogenic activity than suramin [189,190]. In the CAM model (chicken chorioallantoic membrane), the anti-angiogenic result of k-carrageenan oligosaccharides on ECV304 cells was demonstrated to limit cell proliferation, migration, and tube formation [191]. Furthermore, by negatively regulating human bFGFR, bFGF, CD105, and VEGF, oligosaccharides inhibited the formation of new blood vessels in MCF-7 xenograft tumors. Human umbilical vein endothelial cells were treated with λ-carrageenan oligosaccharides at relatively low concentrations (150–300 µg/mL), which had an adverse impact on the development of tumor blood vessel endothelial cells [192]. The amount and position of sulfation, as well as the molecular weight, influence the biological activity of sulfated polysaccharides. Chemical changes, in other words, alter the biological activities of carbohydrates [193]. For example, λ-carrageenan can be broken down into five different compounds with varying molecular weights, all of which have anti-cancer properties, most likely due to immunomodulation. Lower molecular weight products, such as those with molecular weights of 15 and 9.3 kDa, demonstrated superior anti-cancer and immunomodulatory properties [193]. Sulfation, acetylation, and phosphorylation improved the anti-cancer and immunomodulatory properties of k-carrageenan oligosaccharides from Kappaphycus striatum. Chemical modifications increased the oxidant activity of k-carrageenan oligosaccharides as well [194]. Sulfated polysaccharides from red algae and their apoptosis modulation in cancer therapeutics are displayed in Table 3. Induction of apoptosis is the mechanism adopted by chemopreventives. Different sulfated polysaccharides derived from different seaweeds trigger apoptosis in diverse cancer cell lines (Figure 6). Sulfated polysaccharides displayed different chemopreventive roles in cancer (Figure 7).

Figure 6. Apoptosis modulation by different sulfated polysaccharides derived from different sea weeds in cancer prevention.

Figure 7. Sulfated polysaccharides displayed different chemopreventive roles in cancer.
Table 3. Sulfated polysaccharides from red algae and their apoptosis modulating in cancer thera

5. Nanoparticle Synthesis by Using Sulfated Polysaccharides and Its Impact on the Cancer Therapeutic Efficacy
The three main cancer treatments currently available are surgery, chemotherapy, and radiation therapy; chemotherapy, however, has not been the mainstay of cancer care in recent years due to the level to which it can harm healthy normal cells. Nanoparticles have emerged as alternative techniques for addressing only cancer cells, increasing the obtainability of drugs to cancer cells while sparing healthy cells from harm [195]. Seaweeds are a common source of natural sulfated polysaccharides, but there are other sources as well. Numerous biological and biomedical applications have been investigated for Ivan, carrageenan, porphyrin, fucoidan, and their other derivatives in wound management, tissue engineering, drug delivery, and biosensors [196]. Seaweed polysaccharides interact with biological tissue readily because they have hydrophilic surface groups like carboxyl, hydroxyl, and sulfate [197]. Preparatory techniques that produce sulfated polysaccharide nanoparticles with the desired properties for efficient drug delivery systems have received a lot of attention [198,199]. Ionic gelation is typically a straightforward and gentle process for creating sulfated polysaccharide nanoparticles. However, to create ulvan, fucoidan, porphyrin, and carrageenan-based nanoparticles with the desired shape, process optimization is crucial. The optimization can be carried out by adjusting the pH, temperature, concentration of calcium ions, concentration of sulfated polysaccharide, addition speed, and stirring rate. Both MCF7 and HepG2 cells are inhibited from proliferating by van in nanoparticle albumin due to an increase in caspase-8 and caspase-9 levels, which denotes the induction of apoptosis [129]. When creating gold nanoparticles (AuNps), which are used as drug delivery systems for anticancer treatments, porphyrin can also be used as a reducing agent. For instance, a human glioma cell line is more toxic to AuNps coated with porphyrin (LN-229). As a result, porphyrin-capped AuNps were developed and used as doxorubicin hydrochloride anticancer drug carriers [200]. A thymidylate synthase inhibitor called 5-fluorouracil (5-FU) has been used to treat cancer for a long time, but its use has been restricted because of side effects [201]. To create a water-soluble macromolecule for the prodrug 5-FU, porphyrin-capped AuNps can be used as a drug carrier, delaying the 5-FU release and minimizing side effects [202]. Porphyran-capped AuNPs were found to be safe in an in vitro cytotoxicity study, suggesting that they could be used as drug delivery systems [203]. Because of this, using porphyrin as a reducing agent carrier for drug delivery has no unfavorable effects and might make it possible for anticancer medications to work more quickly. Fucoidan porphyran, and carrageenan-based nanoparticles in particular have been thoroughly investigated for the delivery of anti-cancer medications (Table 4).
Table 4. Nanoparticle synthesis by using sulfated polysaccharides and its impact on the cancer therapeutic efficacy.

6. Sulfated Polysaccharides Research Limitations and Future Expansion in Cancer Prevention
Although sulfated polysaccharides have numerous medicinal uses, their low bioavail ability makes them impractical to use in daily life. Different sulfated polysaccharide structures affect how well they are absorbed in different organs [7]. Additionally, a continuous fluctuation in the effective doses in both in vitro and in vivo applications compromises their clinical trial [7]. The in vitro effectiveness of sulfated polysaccharides is frequently not replicated in preclinical or clinical studies [215]. Additionally, their sluggish intracellular metabolism and restricted solubility make clinical application more challenging [216]. More significantly, their wide therapeutic application is a result of their cellular specificity and molecular target selectivity. Depending on the cellular, tissue, and tumor settings, these bioactive chemicals have different ways of causing cell death [216]. Additionally, clinical studies are more successful when the mono-specific and multi-specific functions of action are understood [216]. Synthetic analogs of sulfated polysaccharides might be more bioavailable if they are created and tested [217]. To increase bioavailability and target specificity, sulfated polysaccharides and their synthetic equivalents may benefit from the use of micro-emulsions, nano-carriers, polymers, liposomes, and micelles [218]. These techniques, in our opinion, will be more frequently used in the future to create polysaccharide-based nanoparticles. In terms of delivering anticancer medications with increased bioavailability, seaweed polysaccharide-based nanoparticles have demonstrated promising results [170]. These techniques will also enhance their metabolism in host systems and solubility [170]. Additionally, the preclinical and clinical efficacy of apoptosis will be enhanced by its target specificity. Combining sulfated polysaccharides with drugs that have received FDA approval could significantly increase clinical effectiveness [170]. Additionally, sulfated polysaccharides, when added to or used as adjuvants in food, improve the therapeutic efficacy of modern medications [170].

Benefits of cistanche tubulosa-Antitumor
7. Conclusions and Future Perspectives
The current cancer therapy system has identified sulfated polysaccharides as a trustworthy source for discovering bioactive druggable molecules with a variety of chemotherapeutic effects in various malignancies. Over half of the FDA-approved medications in recent years have been directly extracted from marine sources or created using a chemical counterpart. The isolation and use of these sulfated compounds from marine sources have greater bioavailability, diversified chemical makeup, and non-reductant cytotoxicity. Owing to these characteristics, the seaweed-derived sulfated polysaccharides act as possible lead pharmacophores in treating various malignancies. However, a significant barrier to their pharmaceutical utilization is their bioavailability, improved separation, cleanliness of the isolates, and target selectivity as one drug multi-target specificity and cell/tissue/cancer context. Additionally, they play a significant role as druggable mediators due to their wide variety of therapeutic interventions, low-cost commercial production, and promising pre-clinical and clinical applications. Meanwhile, there is some optimism for commercializing these sulfated polysaccharides from marine seaweeds due to the extensive on- and off-site harvesting of the organisms and low-cost cultivation upkeep. Additionally, the large-scale manufacture of these sulfated polysaccharides for chemotherapy is made more effective by the out-of-range application of chemical synthesis of these polysaccharides. With the advent of new prospects for the isolation and screening of sulfated polysaccharides from seaweed as innovative pharmacological agents against various cancers, the chemotherapeutic use of such prospective agents is likely to flourish shortly. Moreover, nanoparticle-mediated sulfated polysaccharide-based nanoparticles are capable of sustained drug release, high stability, and biocompatibility, all of which will be used in clinical trials in the future. Targeting moieties will increase the therapeutic efficacy of polysaccharide-based nanoparticles while minimizing undesirable side effects. Additionally, creating such drug candidates will improve currently available medications for the advancement of personalized and precision medicine.
References
1. Younossi, Z.M.; Corey, K.E.; Lim, J.K. AGA clinical practice update on lifestyle modification using diet and exercise to achieve weight loss in the management of nonalcoholic fatty liver disease: Expert review. Gastroenterology 2021, 160, 912–918. [CrossRef] [PubMed]
2. Blix, H. Verification of nuclear non-proliferation: Securing the future. IAEA Bull. 1992, 34, 2–5.
3. World Health Organization. Global Status Report on Alcohol and Health 2018; World Health Organization: Geneva, Switzerland, 2019.
4. Edwards, B.K.; Noone, A.M.; Mariotto, A.B.; Simard, E.P.; Boscoe, F.P.; Henley, S.J.; Jemal, A.; Cho, H.; Anderson, R.N.; Kohler, B.A. Annual Report to the Nation on the status of cancer, 1975–2010, featuring prevalence of comorbidity and impact on survival among persons with lung, colorectal, breast, or prostate cancer. Cancer 2014, 120, 1290–1314. [CrossRef] [PubMed]
5. Delgermaa, V.; Takahashi, K.; Park, E.-K.; Le, G.V.; Hara, T.; Sorahan, T. Global mesothelioma deaths reported to the World Health Organization between 1994 and 2008. Bull. World Health Organ. 2011, 89, 716–724. [CrossRef] [PubMed]
6. Pradhan, B.; Nayak, R.; Patra, S.; Jit, B.P.; Ragusa, A. Bioactive Metabolites from Marine Algae as Potent Pharmacophores against Oxidative Stress-Associated Human Diseases: A Comprehensive Review. Molecules 2020, 26, 37. [CrossRef]
7. Pradhan, B.; Patra, S.; Nayak, R.; Behera, C.; Dash, S.R.; Nayak, S.; Sahu, B.B.; Bhutia, S.K.; Jena, M. Multifunctional role of fucoidan, sulfated polysaccharides in human health and disease: A journey under the sea in pursuit of potent therapeutic agents. Int. J. Biol. Macromol. 2020, 164, 4263–4278. [CrossRef]
8. Patra, S.; Bhol, C.S.; Panigrahi, D.P.; Praharaj, P.P.; Pradhan, B.; Jena, M.; Bhutia, S.K. Gamma irradiation promotes chemo sensitization potential of gallic acid through attenuation of autophagic flux to trigger apoptosis in an NRF2 inactivation signaling pathway. Free Radic. Biol. Med. 2020, 160, 111–124. [CrossRef]
9. Srivastava, A.; Rikhari, D.; Pradhan, B.; Bharadwaj, K.K.; Gaballo, A.; Quarta, A.; Jena, M.; Srivastava, S.; Ragusa, A. An Insight into Neuropeptides Inhibitors in the Biology of Colorectal Cancer: Opportunity and Translational Perspectives. Appl. Sci. 2022, 12, 8990. [CrossRef]
10. Gutiérrez-Rodríguez, A.G.; Juárez-Portilla, C.; Olivares-Bañuelos, T.; Zepeda, R.C. Anticancer activity of seaweeds. Drug Discov. Today 2018, 23, 434–447. [CrossRef]
11. Jit, B.P.; Pattnaik, S.; Arya, R.; Dash, R.; Sahoo, S.S.; Pradhan, B.; Bhuyan, P.P.; Behera, P.K.; Jena, M.; Sharma, A.; et al. Phytochemicals: A potential next-generation agent for radioprotection. Phytomed. Int. J. Phytother. Phytopharm. 2022, 2022, 154188. [CrossRef] [PubMed]
12. Jit, B.P.; Pradhan, B.; Dash, R.; Bhuyan, P.P.; Behera, C.; Behera, R.K.; Sharma, A.; Alcaraz, M.; Jena, M. Phytochemicals: Potential Therapeutic Modulators of Radiation-Induced Signaling Pathways. Antioxidants 2022, 11, 49. [CrossRef] [PubMed]
13. Panigrahi, G.K.; Yadav, A.; Mandal, P.; Tripathi, A.; Das, M. Immunomodulatory potential of rhein, an anthraquinone moiety of Cassia occidentalis seeds. Toxicol. Lett. 2016, 245, 15–23. [CrossRef] [PubMed]
14. Ovadje, P.; Roma, A.; Steckle, M.; Nicoletti, L.; Arnason, J.T.; Pandey, S. Advances in the research and development of natural health products as mainstream cancer therapeutics. Evid. Based Complement. Altern. Med. 2015, 2015, 751348. [CrossRef] [PubMed]
15. Patra, S.; Nayak, R.; Patro, S.; Pradhan, B.; Sahu, B.; Behera, C.; Bhutia, S.K.; Jena, M. Chemical diversity of dietary phytochemicals and their mode of chemoprevention. Biotechnol. Rep. (Amst. Neth.) 2021, 30, e00633. [CrossRef]
16. Patra, S.; Pradhan, B.; Nayak, R.; Behera, C.; Das, S.; Patra, S.K.; Efferth, T.; Jena, M.; Bhutia, S.K. Dietary polyphenols in chemoprevention and synergistic effect in cancer: Clinical evidence and molecular mechanisms of action. Phytomed. Int. J. Phytother. Phytopharm. 2021, 90, 153554. [CrossRef] [PubMed]
17. Patra, S.; Pradhan, B.; Nayak, R.; Behera, C.; Panda, K.C.; Das, S.; Jena, M. Apoptosis and autophagy modulating dietary phytochemicals in cancer therapeutics: Current evidence and future perspectives. Phytother. Res. 2021, 35, 4194–4214. [CrossRef]
18. Patra, S.; Pradhan, B.; Nayak, R.; Behera, C.; Rout, L.; Jena, M.; Efferth, T.; Bhutia, S.K. Chemotherapeutic efficacy of curcumin and resveratrol against cancer: Chemoprevention, chemoprotection, drug synergism and clinical pharmacokinetics. In Proceedings of the Seminars in Cancer Biology; Academic Press: Cambridge, MA, USA, 2021; pp. 310–320.
19. Wilson, R.M.; Danishefsky, S.J. Small molecule natural products in the discovery of therapeutic agents: The synthesis connection. J. Org. Chem. 2006, 71, 8329–8351. [CrossRef] [PubMed]
20. Simmons, T.L.; Andrianasolo, E.; McPhail, K.; Flatt, P.; Gerwick, W.H. Marine natural products as anticancer drugs. Mol. Cancer Ther. 2005, 4, 333–342. [CrossRef]
21. Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2019, 36, 122–173. [CrossRef] [PubMed]
22. Shinde, P.; Banerjee, P.; Mandhare, A. Marine natural products as a source of new drugs: A patent review (2015–2018). Expert Opin. Ther. Pat. 2019, 29, 283–309. [CrossRef] 23. Pradhan, B.; Kim, H.; Abbasi, S.; Ki, J.-S. Toxic Effects and Tumor Promotion Activity of Marine Phytoplankton Toxins: A Review. Toxins 2022, 14, 397. [CrossRef]
24. Pradhan, B.; Nayak, R.; Bhuyan, P.P.; Patra, S.; Behera, C.; Sahoo, S.; Ki, J.-S.; Quarta, A.; Ragusa, A.; Jena, M. Algal Phlorotannins as Novel Antibacterial Agents concerning the Antioxidant Modulation: Current Advances and Future Directions. Mar. Drugs 2022, 20, 403. [CrossRef] [PubMed]
25. Pradhan, B.; Ki, J.-S. Phytoplankton Toxins and Their Potential Therapeutic Applications: A Journey toward the Quest for Potent Pharmaceuticals. Mar. Drugs 2022, 20, 271. [CrossRef] [PubMed]
26. Pradhan, B.; Maharana, S.; Bhakta, S.; Jena, M. Marine phytoplankton diversity of Odisha coast, India with special reference to new record of diatoms and dinoflagellates. Vegetos 2021, 35, 330–344. [CrossRef]
27. Behera, C.; Dash, S.R.; Pradhan, B.; Jena, M.; Adhikary, S.P. Algal Diversity of Ansupa lake, Odisha, India. Nelumbo 2020, 62, 207–220. [CrossRef]
28. Behera, C.; Pradhan, B.; Panda, R.; Nayak, R.; Nayak, S.; Jena, M. Algal Diversity of Saltpans, Huma (Ganjam), India. J. Indian Bot. Soc. 2021, 101, 107–120. [CrossRef]
29. Dash, S.; Pradhan, B.; Behera, C.; Jena, M. Algal Diversity of Kanjiahata Lake, Nandankanan, Odisha, India. J. Indian Bot. Soc. 2020, 99, 11–24. [CrossRef]
30. Dash, S.; Pradhan, B.; Behera, C.; Nayak, R.; Jena, M. Algal Flora of Tampara Lake, Chhatrapur, Odisha, India. J. Indian Bot. Soc. 2021, 101, 1–15. [CrossRef]
31. Maharana, S.; Pradhan, B.; Jena, M.; Misra, M.K. Diversity of Phytoplankton in Chilika Lagoon, Odisha, India. Environ. Ecol 2019, 37, 737–746.
32. Mohanty, S.; Pradhan, B.; Patra, S.; Behera, C.; Nayak, R.; Jena, M. Screening for nutritive bioactive compounds in some algal strains isolated from coastal Odisha. J. Adv. Plant Sci. 2020, 10, 1–8.
33. Pradhan, B.; Patra, S.; Dash, S.R.; Satapathy, Y.; Nayak, S.; Mandal, A.K.; Jena, M. In vitro, antidiabetic, anti-inflammatory, and antibacterial activity of marine alga Enteromorpha compresses collected from Chilika lagoon, Odisha, India. Vegetos 2022, 35, 614–621. [CrossRef]
34. Pradhan, B.; Nayak, R.; Patra, S.; Bhuyan, P.P.; Behera, P.K.; Mandal, A.K.; Behera, C.; Ki, J.-S.; Adhikary, S.P.; Mubarak Ali, D.; et al. A state-of-the-art review on fucoidan as an antiviral agent to combat viral infections. Carbohydr. Polym. 2022, 2022, 119551. [CrossRef] [PubMed]
35. Pradhan, B.; Nayak, R.; Patra, S.; Bhuyan, P.P.; Dash, S.R.; Ki, J.-S.; Adhikary, S.P.; Ragusa, A.; Jena, M. Cyanobacteria and Algae-Derived Bioactive Metabolites as Antiviral Agents: Evidence, Mode of Action, and Scope for Further Expansion; A Comprehensive Review in Light of the SARS-CoV-2 Outbreak. Antioxidants 2022, 11, 354. [CrossRef] [PubMed]
36. Pradhan, B.; Patra, S.; Dash, S.R.; Nayak, R.; Behera, C.; Jena, M. Evaluation of the anti-bacterial activity of methanolic extract of Chlorella vulgaris Beyerinck [Beijerinck] with special reference to antioxidant modulation. Future J. Pharm. Sci. 2021, 7, 17. [CrossRef]
37. Pradhan, B.; Patra, S.; Behera, C.; Nayak, R.; Jit, B.P.; Ragusa, A. Preliminary Investigation of the Antioxidant, Anti-Diabetic, and Anti-Inflammatory Activity of Enteromorpha intestina lis Extracts. Molecules 2021, 26, 1171. [CrossRef]
38. Pradhan, B.; Patra, S.; Behera, C.; Nayak, R.; Patil, S.; Bhutia, S.K.; Jena, M. Enteromorpha compress an extract induces anticancer activity through apoptosis and autophagy in oral cancer. Mol. Biol. Rep. 2020, 47, 9567–9578. [CrossRef]
39. Cumashi, A.; Ushakova, N.A.; Preobrazhenskaya, M.E.; D’Incecco, A.; Piccoli, A.; Totani, L.; Tinari, N.; Morozevich, G.E.; Berman, A.E.; Bilan, M.I. A comparative study of the anti-inflammatory, anticoagulant, antiangiogenic, and antiadhesive activities of nine different fucoidans from brown seaweeds. Glycobiology 2007, 17, 541–552. [CrossRef]
40. Park, H.Y.; Han, M.H.; Park, C.; Jin, C.-Y.; Kim, G.-Y.; Choi, I.-W.; Kim, N.D.; Nam, T.-J.; Kwon, T.K.; Choi, Y.H. Anti-inflammatory effects of fucoidan through inhibition of NF-κB, MAPK and Akt activation in lipopolysaccharide-induced BV2 microglia cells. Food Chem. Toxicol. 2011, 49, 1745–1752. [CrossRef]





