Lipidomics: New Insight Into Kidney Disease
Mar 25, 2022
Contact: Audrey Hu Whatsapp/hp: 0086 13880143964 Email: audrey.hu@wecistanche.com
Ying-Yong Zhao, Nosratola D. Vaziri, Rui-Chao Lin
Abstract
Due to the incidence of type-2 diabetes and hypertension, chronic kidney disease (CKD) has emerged as a major public health problem worldwide. CKD results in premature death from accelerated cardiovascular disease and various other complications. Early detection, careful monitoring of renal function, and response to therapeutic intervention are critical for the prevention of CKD progression and its complications. Unfortunately, traditional biomarkers of renal function are insufficiently sensitive or specific to detect early stages of disease when therapeutic intervention is most effective. Therefore, more sensitive biomarkers of kidney disease are needed for early diagnosis, monitoring, and effective treatment. CKD results in profound changes in lipid and lipoprotein metabolism that, in turn, contribute to the progression of CKD and its cardiovascular complications. Lipids and lipid-derived metabolites play diverse and critically important roles in the structure and function of cells, tissues, and biofluids. Lipidomics is a branch of metabolomics, which encompasses the global study of lipids and their biological function in health and disease including identification of biomarkers for diagnosis, prognosis, prevention, and therapeutic response for various diseases. This review summarizes recent developments in lipidomics and its application to various kidney diseases including chronic glomerulonephritis, IgA nephropathy, chronic renal failure, renal cell carcinoma, diabetic nephropathy, and acute renal failure in clinical and experimental research. Analytical technologies, data analysis, as well as currently known metabolic biomarkers of kidney diseases are addressed. Future perspectives and potential limitations of lipidomics are discussed.

Cistanche deserticola prevents kidney disease, click here to get the sample
1. INTRODUCTION
Due to the incidence of type-2 diabetes and hypertension, chronic kidney disease (CKD) has emerged as a major public health issue worldwide. CKD results in disability and premature death from accelerated cardiovascular disease and its attendant complications [1]. Numerous pathologic conditions include genetic, metabolic, toxic, immunologic, infectious, hemodynamic, mechanical, and other disorders that lead to the development and progression of kidney disease. Early detection, careful monitoring of renal function, and response to therapeutic intervention are critical for timely diagnosis and prevention of progression of CKD and its complications. Unfortunately, traditional markers of renal function are insufficiently sensitive or specific to detect CKD and its cardiovascular or other complications at an early stage when therapeutic intervention is most effective. For example, the most commonly used biomarkers, i.e., serum creatinine and urea and creatinine clearance, are heavily influenced by factors independent of intrinsic kidney function and structure. In this context, muscle mass significantly affects creatinine, protein intake and fluid balance modulate urea, and the use of angiotensin-converting enzyme inhibitors or angiotensin receptor blockers, as well as dietary protein intake, affects creatinine clearance. It is, therefore, necessary to develop sensitive and specific biomarkers for early detection of kidney disease and monitoring of its progression and response to therapeutic intervention. Insight into the dynamic differences in genetic, protein, and metabolite regulation, interaction, and function in kidney diseases might identify new diagnostic and prognostic biomarkers and therapeutic targets [2–4].
CKD results in profound changes in lipid and lipoprotein metabolism [5–7]. The associated lipid disorders, in turn, contribute to the progression of CKD and its cardiovascular and other complications [8–10]. Lipidomics, the global study of lipids within cells, tissues, and biofluids, involves the analysis of lipid species and their abundance to elucidate biologic function, subcellular localization, and tissue distribution. Small molecular weight lipids such as fatty acids, glycerolipids, glycerophospholipids (GPs), and sphingolipids serve diverse and complex functions in health and disease. They play important roles in the regulation of normal kidney function and the pathogenesis of kidney disease. Previous studies have shown significantly increased glomerular cyclooxygenase-1 or -2 expression in inpatient and animal models of glomerulonephritis [11–13] and upregulation of glomerular cyclooxygenase-2 expression in inpatient and animal models of lupus nephritis [13,14]. Cyclooxygenase inhibition has been shown to ameliorate passive Heymann nephritis and lupus nephritis in experimental animals [14–16]. Leukotrienes, associated with inflammatory glomerular injury and lipoxygenase product (12-hydroxyeicosatetraenoic acid), mediated angiotensin II and transforming growth factor-β-induced mesangial expansion in diabetic nephropathy (DN) [17]. 20-Hydroxyeicosatetraenoic and epoxyeicosatrienoic acids were involved in several forms of kidney injury, including renal injury in metabolic syndrome [18–20] and ceramides have been shown to play a role in the pathogenesis of acute kidney injury. Taken together there is mounting evidence supporting the role of lipids and lipid-derived metabolites in the pathogenesis of kidney disease. Thus, analysis of key lipid mediators has emerged as an important tool in the diagnosis, prognosis, and treatment of kidney disease.
This article reviews recent advances in the use of lipidomics in elucidating the pathogenesis and potential treatment of kidney disease.
2. KIDNEY DISEASE
Systems biology allows on-time analysis of regulatory and biologic networks in cell metabolism [21–23]. Comprehensive characterization of renal diseases could provide important and integrative information to better characterize molecular relationships underlying this pathophysiology in order to develop more reliable and specific markers for diagnosis, prognosis, prevention, and therapeutic response [2,24]. The growth of systems biology and the development of new experimental and computational tools have enabled the connection of gene–cell–organ regulatory mechanisms at multiple levels to integrate the molecular and cell biology of kidney structure and function [25–29]. Lipids play diverse and important roles in biologic systems including membrane bilayer structure, energy storage, signal transduction, and also provide functional support for membrane proteins and their interactions [30]. For example, arachidonic acid is the precursor of eicosanoids, which act as signaling molecules via specific receptors leading to inflammatory processes [31]. Triacylglycerides serve as cellular energy storage and play an important role in metabolism and disease [32]. Some lipid species, i.e., lysophosphatidylcholines (LPC), glycerophosphoethanolamines (PE), phosphatidylcholines (PC), and glycerophosphoinositols (PI), appear to be potential kidney disease markers [33]. Here, we provide an overview of the lipidomic approach in kidney disease.

benefit of cistanche: treating kidney diseases
3.LIPIDS AND LIPIDOMICS
3.1.Definition, classification, and biologic function of lipids
Lipids, the fundamental components of biologic membranes, are a structurally and functionally diverse class of molecules. Depending on biosynthesis and chemical structure, lipids are defined as hydrophobic or amphiphilic. Amphiphilic lipids exist in vesicles, membranes, or liposomes in an aqueous environment. Biologic lipids originate two distinct types of biochemical subunits: isoprene and ketoacyl groups [34]. Based on this definition, lipids can be divided into eight categories: fatty acids, glycerolipids, sphingolipids, GPs, saccharolipids, sterol lipids, prenol lipids, and polyketides (Fig. 1) [34]. Fatty acids and glycerolipids have relatively simple structures. Fatty acids are one of the most important lipid classes and basic components of all lipids. Fatty acids have saturated or unsaturated straight-carbon chains with lengths of 4–24 carbon atoms and 0–6 double bonds. Fatty acids are precursors of various bioactive lipids. Eicosanoids include leukotrienes, prostaglandins, and thromboxanes which play important roles in the development of inflammatory processes [35]. Glycerolipids are composed of mono-, di-, and tri-substituted glycerols that differ in fatty acid content esterified to the hydroxyl groups of the glycerol backbone [36]. A variety of studies have shown that altered triglyceride synthesis and catabolism play important roles in the occurrence and development of many diseases [37,38]. Sterol lipids, including cholesterol and their derivatives composed of a fused four-ring core structure, are important components of membrane lipids. Sterol lipids have different biologic roles such as the regulatory function of cell signaling and modulation of cellular fluid [39].
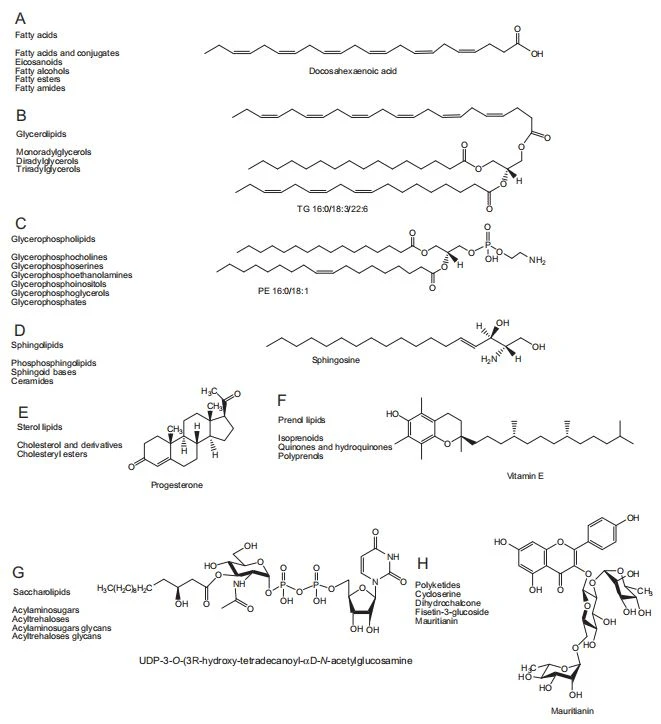
GP, also known as phospholipids, are ubiquitous in nature are important components of lipid bilayers, and are involved with cell signaling and metabolism. Based on the nature of the polar head group at the sn-3 position of the glycerol backbone in eukaryotes and eubacteria or the sn-1 position in the case of archaebacteria [40], GP may be subdivided into distinct classes including glycerophosphocholines, glycerophosphatidic acids, glycerophosphoglycerols (PG), glycerophosphoserines (PS), PE, and PI. Brain tissue contains relatively high G, and alterations of their composition have been implicated in neurologic disorders [41]. Some GP such as LPC, PC, PE, and PI have been identified as potential biomarkers of cancer, kidney, and cardiovascular disease [33,42,43]. Sphingolipids consist of a complex family of compounds composed of a basic backbone of 1,3- dihydroxyl, 2-amino alkane, or alkene (sphingoid base). Sphingomyelin (SM) and sphingosine are two important sphingolipids composed of a phosphorylcholine head group and fatty acid linked to a 1-hydroxyl group and 2-amino group of the sphingoid chain, respectively. Earlier studies have demonstrated that ceramides, which belong to N-acyl-derivatives of sphingosine, are associated with CKD [44].
3.2.Lipidomics
Although a subfraction of the metabolome, its complexity of lipid species, their distinct chemical properties, and important biologic activity have made lipidome the focus of considerable research. Metabolomics is defined as “the quantitative measurement of the dynamic multiparametric metabolic response of living systems to pathophysiologic stimuli or genetic modification” [45,46]. Metabolomics is a nontargeted quantitative analysis of biofluids and tissues for low molecular mass endogenous metabolites. Lipidomics, as a branch of metabolomics, was first introduced by Han and Gross in 2003 [47]. Lipidomics has been defined as “the full characterization of lipid molecular species and of their biological roles in the expression of proteins involved in lipid metabolism and function, including gene regulation” [48]. Lipidomics represents a shift from individual lipid study to the examination of global lipid metabolites in a systems-integrated context to more fully understand their role in pathophysiologic processes. In the past 10 years, lipidomics has emerged as a new field in systems biology and has increased interest in disease diagnosis and biomarker discovery (obesity, diabetes, cardiovascular disease, Alzheimer’s disease, pancreatic cancer, etc.), pharmaceutical discovery and development, human food and nutrition research [49–55]. This powerful approach can reveal unique metabolic characteristics of normal, pathologic, or treatment-specific events. Recently, an increased number of lipidomic studies and reviews have been published using mass spectrometry (MS), nuclear magnetic resonance (NMR), and other spectroscopic modalities [56–61]. Separation technologies, gas chromatography (GC), liquid chromatography (LC), supercritical fluid chromatography, and capillary electrophoresis, are critical for the lipidomic examination of complex samples [62]. To obtain structural molecular ion information, low collision energy MS is first used followed by higher collision energy MS2 conditions to obtain fragment ions. Typically the precursor ion is selected and fragmentation monitored by tandem mass spectrometry (MS/MS). This approach provides greater structural information and detection of individual lipid species in complex biological samples. In addition, MS/MS has been increasingly used to develop quantitative methods for targeted lipidomics [63]. This approach, however, requires information based on a previous full-scan MS. In 2005, Wrona et al.[64] introduced the MSE technique in which two scanning functions are simultaneous for data collection. MSE provided parallel alternating scans for acquisition at low collision energy for precursor ion information (MS) or high collision energy for full-scan accurate mass fragments, precursor ions, and neutral loss information (MSE). This approach provided similar information to conventional MS2 (MS/MS) in a single analytical run and structural information required for the identification of unknown biomarkers in untargeted analyses [65–70]

cistanche health benefits
3.3.Analytical methods for lipidomics
The traditional methods of lipid analysis usually involve solvent extraction of the biologic samples (blood, tissue, cell, and organism) followed by separation of lipids using thin-layer chromatography, solid-phase extraction, or normal-phase LC and separation of particular classes of lipids into individual molecular species by high-performance liquid chromatography (HPLC)- ultraviolet detector or evaporative light-scattering detector. Using these traditional methods, single molecular species of many lipid classes can be analyzed [71]. Although GC has been used to determine the fatty acid content of the various lipids by the methyl esters method, this approach tends to be time-consuming and involves specimen hydrolysis and derivatization. In general, conventional lipid analysis typically requires a large sample quantity because many biologically active species are present in very small amounts. Due to their inherent complexity, sample preparation may involve multiple extractions thus further lowering sensitivity and resolution. In addition, these methods are labor-intensive and often require derivatization thus limiting throughput.
In contrast, direct sample analysis may be used for MS lipidomics [72,73]. Direct-infusion MS technologies have good reproducibility, accuracy, and high sensitivity and are less time-consuming than traditional methods. Typically, electrospray ionization quadrupole-time-of-flight (ESI-QTOF) and matrix-assisted laser desorption ionization (MALDI) are the most widely used ion sources in direct-infusion MS analysis [74,75]. Direct-infusion MS is simple and fast. Its main limitation is ion suppression, which hampers sensitivity and quantitative accuracy. Unfortunately, this method is unable to identify isobaric and isomeric lipids, whose masses are identical and often produce similar fragmentation patterns. Although direct-infusion MS is relatively limited in searching for novel and unknown compounds from lipid databases, it may be useful to screen biochemical pathways in various diseases in the future. Comprehensive reviews of direct-infusion ESI/ MS, ESI-QTOF/MS, and MALDI/MS and their applications in lipidomics have been published [74,75].
MS is commonly combined with LC for lipidomics and LC–MS-based studies in lipidomics have been reviewed [76]. Typically, the advantages of the LC-MS approach are good reproducibility, accuracy, and high sensitivity for the identification of known or novel lipids. Over the past decade, HPLC–MS has been widely used for both targeted and untargeted analyses in metabolomics and lipidomics using single quadrupole, hybrid, and high-resolution instruments. For global profiling, combinations of ultra-performance liquid chromatography (UPLC) coupled with QTOF/MS or tandem ion mobility TOF/MS are popular choices [77–80]. These provide rapid analysis with high-resolution MS. UPLC uses sub-2 μm sized particles and operates at increased pressures (6000–15,000 psi) thus providing high chromatographic resolution versus conventional HPLC with 5 μm particles [81]. Increased resolution results from improved signal/noise ratio and narrow peak width versus conventional HPLC. This approach is advantageous for metabolic profiling because an enormous number of metabolites may be detected at physiologic concentrations. Although lipids from various biologic sources may be separated by UPLC–MS [82], matrix effects have an important influence on global profiles [83]. Unfortunately, sensitivity is typically not as high as targeted lipidomics. In addition, the experimental conditions of each separated compound cannot be optimized. Typically, triple quadrupole MS is used for targeted analyses by UPLC–MS with selective ion monitoring. Targeted lipid methods may include sterols and eicosanoids such as bile acids and steroids [84,85]. GC-based methods are suitable for volatile components and cannot be used for most lipids. Interestingly, GC–MS is the most widely used method for the analysis of free fatty acids, esterified fatty acids, and steroids. Free fatty acids and steroids require derivatization or silylation, whereas esterified fatty acids are often analyzed as methyl esters [86]. Supercritical fluid chromatography is another high-resolution technique that can be used for the separation of various lipids. Supercritical fluid chromatography MS can be used for comprehensive lipid profiling of large sample numbers [87].
Ion mobility MS (IM-MS) and multidimensional methodologies are regarded as novel methodologies and have been used in lipidomics [88,89]. Isomers, conformers, and enantiomers can be rapidly separated by IM-MS and have proven useful in the analysis of complex biological samples [78]. The development of imaging MS has also played an important role in the development of imaging ion mobility spectrometry with MS for lipid analysis. Ion mobility spectrometry with MS combined with molecular dynamics computational modeling can use for future characterization of the structure and stability of lipid-incorporated complexes. In addition, comprehensive multidimensional LC-MS is an attractive emerging approach for comprehensive lipidomic characterization of complex biologic samples [90].
3.4.Analysis of lipidomics data
Lipidomics produces enormous data and its analysis play a key role, especially in untargeted studies. As such, robust bioinformatics is critical. Prior to statistical analysis, data preprocessing including signal processing, data normalization, and transformation are required, such that raw data are transformed into a format compatible with statistical data analysis [91,92]. Given the large degree of lipid variation, the first step of unsupervised and supervised statistical analysis is data reduction. This may be accomplished by a number of methods including orthogonal partial least squares-discriminate analysis, principal components analysis (PCA), and partial least squares- discriminate analysis (PLS-DA). Both unsupervised and supervised methods can be used, depending on the goal of the specific analysis. In unsupervised data analysis, unknown information about different groups is used by PCA and hierarchical cluster analysis. In the supervised approach, each sample or metabolite is associated with known compounds, and this prior information is then used for analysis via principal component regression and neural networks [91,92]. Other regression methods including Elastic Net and Least Absolute Shrinkage and Selection Operator are also available for analysis of lipidomic data sets to ascertain the relationship between variables [93].

cistanche testosterone: improve kidney function
4. APPLICATIONS OF LIPIDOMICS IN KIDNEY DISEASES
Phospholipids represent a class of important cellular constituents that participate in numerous biologic processes and pathways reflective of metabolic status in health and disease. Lipidomics is a suitable tool for disease biomarker discovery in systems biology [94,95]. A comprehensive understanding of its applications is critically important for lipidomics. Many studies have demonstrated that metabolic disorders or abnormalities of various lipids lead to kidney disease [96–99]. Using chronic renal failure (CRF), renal cell carcinoma (RCC), chronic glomerulonephritis, IgA nephropathy, and DN, we discuss lipidomics in kidney disease in humans and animals- and cell-model studies.
4.1.Lipidomics in clinical kidney disease
4.1.1Effect of chronic kidney disease and glomerulonephritis
Lipid abnormalities are common in kidney disease [100,101] and contribute to a high incidence of cardiovascular disorders in this population. Plasma and erythrocyte lipid profiles were examined in CRF patients on hemodialysis for 30 months [102]. Increased triglyceride was observed in plasma and erythrocyte membranes. Increased plasma palmitic acids and monounsaturated fatty acids and decreased plasma polyunsaturated fatty acids were also observed in CRF. Lipid abnormalities were apparent at 18 months and became more profound at 30 months. Plasma and erythrocyte membrane lipid patterns did not change during the dialysis period. CRF patients under regular hemodialysis showed a gradual deterioration in the triglyceride and fatty acid profiles. In another study, HPLC–MS was used to profile plasma phospholipids in patients with chronic glomerulonephritis and CRF without renal replacement therapy [103]. The results showed that primary chronic glomerulonephritis and CRF had abnormal metabolic phospholipid profiles. A number of phospholipids (n ¼ 19) were identified as potential biomarkers. A possible mechanism leading to this abnormality included phosphatidylinositol (PI) hydrolysis via activation of PI-specific phospholipase C, leading to the production of two-second messengers, inositol (1,4,5)- trisphosphate (IP3) and diacylglycerol [104], that participate in signal transduction independently. IP3 increases cytoplasmic Ca2+ by stimulating the release of Ca2+ from the sarcoplasmic reticulum [105]. Protein kinase C (PKC) is activated by phosphatidylserine, Ca2+, and diacylglycerol. Activation of the intracellular PKC signal transduction system, in turn, triggers a series of physiologic and physiochemical reactions.
Based on morphologic and genetic features, RCC is classified into various subtypes. RCC prognosis varies and metastatic or recurrent RCC is associated with poor prognosis with rare long-term survival. Desorption ESI/MS was used in an imaging mode to study the lipid profiling of thin tissue sections of human papillary RCC versus adjacent normal tissue (11 sample pairs) and clear cell RCC versus adjacent normal tissue (9 sample pairs) [106]. Increased GP and free fatty acids were observed in the tumor region. PLS- DA distinguished tumor in papillary and clear cell RCC and papillary from clear cell RCC. Altered GP tissue composition occurs in cancer [107] and appears integrally associated with malignant transformation [108]. Micro- LC-QTOF/MS was used to investigate urine lipids in RCC versus healthy subjects. Thirty-five lipid species were tentatively identified including lipidomic changes in urinary exosomes [109]. Tissue GP and their enzymatic by-products appear related to malignant transformation [110,111] and significantly increased PI has been observed in transformed cells [112].
4.1.2Effect of DN
DN is a serious problem worldwide. Phospholipids and their metabolites are closely related to the pathogenesis and progression of DN. Untargeted lipidomics of serum phospholipids using normal-phase LC-TOF/MS and ion trap-MS/MS was performed on DN patients [113]. Comparison with healthy subjects revealed eight lipids in seven phospholipid classes as potential DN biomarkers. Two novel biomarkers including PI (18:0/22:6) and SM (d18:0/20:2) effectively discriminated against DN patients. Predictably, the same phospholipid class has a similar variation trend with DN progression. Upregulated LPC, PE, PG, SM, one PC, and one PI and downregulated PE, PS, and two PC were noted. A number of studies have shown lipid accumulation in the kidneys of diabetic experimental animals and humans and that lipids influenced the DN pathogenesis [114,115]. It was reported that lipid phosphatase promoted podocyte apoptosis leading to DN and lipid phosphatase was increased before histologic change [116]. Additional evidence has shown that abnormal lipid metabolism and accumulation of lipids in the kidney played an important role in DN pathogenesis [117–119] and oxidized PC species were related to renal dysfunction [120]. Possible mechanisms involve lipid deposition due to increased serum concentration as well as glomerular filtration of protein-bound lipids associated with proteinuria. Accumulated lipids increased the expression of vascular endothelial growth factors and transformed growth factor-β as well as promotion of proteinuria and diabetic glomerulosclerosis [121]. On the other hand, the presence of abnormal phospholipids may promote activation of the sorbitol pathway, oxidative stress, and activation of PKC [122–124]. In DN, decreased PI was related to activation of the sorbitol pathway leading to degradation of intra- cellular inositol, reduction of myoinositol, and reduction of PI synthesis.
4.1.3Effects of renal replacement modalities
Clinical complications associated with peritoneal dialysis have become increasingly evident. An online two-dimensional LC-QTOF/MS was developed for plasma lipid profiling in peritoneal dialysis patients [125]. This comprehensive study included 10 lipid classes and 190 lipid species. Thirty biomarkers were identified including PE and PC as indicators of malnutrition, inflammation, and atherosclerotic syndrome. This study also examined differences in lipid profiles in plasma from individuals with poor fluid control and those with good volume status. Significantly increased PC and PE (and plasmalogen subclasses of PC and PE) were observed in those with poor volume status. Interestingly, another similar study showed that the incidence of malnutrition was associated with plasmalogen phospholipids [126]. These findings supported an association between volume and nutritional status in peritoneal dialysis [127]. GC–MS was used to quantify F2-isoprostanes in hemodialysis patients with end-stage renal disease [128]. F2-isoprostanes were increased ~100-fold after iron/ascorbate-induced oxidative stress and 2- to 4-fold after pentylenetetrazol-induced seizures in hemodialysis patients. Both human and experimental studies support an association between F2-isoprostanes and inflammation.

cistanche dosage
4.2.Lipidomics in animal models or cell models
4.2.1Effect of IgA nephropathy
IgA nephropathy is the most common form of glomerulonephritis and can progress to end-stage renal failure. To identify markers of progression, HPLC–MS with PCA and PLS-DA was used to evaluate phospholipid metabolic profiles in plasma in an experimental Balb/c mouse model [129]. PC, LPC, PI, PS, PE, and SM lipid classes including 90 lipid species were identified. PS(18:0/18:0), PS(18:0/22:5), and PI(18:0/20:4) were identified as potential biomarkers. The relationship of phospholipids and intercellular adhesion molecule-1 (ICAM-1) expression was also examined. The latter is highly correlated to proteinuria. Another study identified ICAM-1 expression as an indicator of disease progression and suggested PS(18:0/18:0), PS(18:0/22:5), and PI(18:0/20:4) as possible biomarkers of IgA nephropathy [130].
Imaging MS lipidomics is useful for visualizing the localization of various lipids in the kidney and other tissue [131,132]. Recently the molecular distribution of lipids was analyzed in hyper-IgA murine kidneys using MALDI-quadrupole ion trap-TOF-based imaging MS [133]. Two PC, PC(18:2/22:6) and PC(16:0/22:6) were primarily found in the cortex and two triacylglycerols, TAG(18:1/18:2/18:1) and TAG(16:0/18:2/18:1), were found in the hilum. However, several other lipids were observed in hyper-IgA kidneys, especially in the tubular region. Two hyper-IgA-specific lipids were O-PC including PC(O-18:1/22:6) and PC(O-16:0/22:6). It was reported that PC(O-18:1/22:6) and PC(O-16:0/22:6) were analogs of plasmalogen and platelet-activating factor, respectively [134,135]. This study also indicated that all the hyper-IgA-specific lipids were derived from urine and that stagnation due to unilateral ureteral obstruction caused the hyper-IgA-specific distribution of lipids in renal tubules.
A possible mechanism involved activation of the PKC pathway leading to expansion of extracellular matrix and thickening of glomerular basement membrane [136]. In fact, PKC activation has been shown to increase the permeability of the endothelial monolayer to albumin [137]. Epithelial cells and the basement membrane from the glomerular capillary barrier. PKC activation has been shown to damage the glomerular capillary barrier leading to proteinuria [138,139].
4.2.2Effect of DN
Rapamycin was shown to prevent DN development in streptozotocin-induced diabetic rats. MALDI-TOF/MS of the renal cortex revealed three classes of sphingolipids including ceramides, SM, and ceramide mono- hexoses [140]. One ceramide metabolite was significantly increased whereas three disappeared. Sphingolipid composition was greatly altered by rapamycin treatment. Increased ceramide(d18:0/16:0), ceramide mono hexoside(d18:1/15:0), SM(d16:1/18:0), and SM(d18:1/18:0) were reversed by rapamycin. The previous study showed that ceramide increase in the diabetic kidney and decrease after rapamycin treatment and the long-established relationship of ceramide and apoptosis support ceramide as a reasonable biomarker candidate [141]. Streptozotocin significantly increased the synthesis of many sphingolipids that was inhibited by rapamycin. Other studies showed that ceramide inhibition, via blocking ceramide synthase or serine palmitoyltransferase, effectively decreased cell death caused by hypoxia-reoxygenation, chemical hypoxia, and radiocontrast media in renal tubular epithelial cells [142–144].
4.2.3Effect of acute renal failure
Inflammation plays a key role in the pathogenesis of acute renal failure [145,146]. LC-MS lipidomics was used to investigate the impact of short-term dietary ω-3 or ω-6 polyunsaturated fatty acids on ischemic renal injury and renal lipid autacoid circuits [147]. Renal ischemia (30 min) resulted in significantly reduced renal function and significantly increased serum creatinine in mice fed a ω-6 supplemented diet but remained normal in mice fed a ω-3 supplemented diet. Moreover, an extension of renal ischemia (45 min) caused 100% mortality in ω-6 supplemented mice but no death in the ω-3 supplemented group. The protective effect of ω-3 polyunsaturated fatty acids against ischemic renal injury was associated with decreased polymorphonuclear leukocyte recruitment, chemokine, and cytokine production, abrogated formation of lipoxygenase- and cyclooxygenase-derived eicosanoids, and increased protectin D1 expression [148]. Systemic treatment with protectin D1 decreased kidney polymorphonuclear leukocyte influx and upregulated the expression of heme oxygenase-1 protein and mRNA in injured and uninjured kidneys. Protectin D1 appeared effective in the prevention of acute kidney injury as well as the effect of dietary ω-3 and ω-6 polyunsaturated fatty acids on autacoid formation in the kidney and outcome of ischemic renal injury [149].
4.2.4Cell research
ESI/MS lipidomics was used to identify phospholipid changes in the human embryonic kidney (HEK293) and human kidney carcinomas (Caki-1) cell death [150]. Significantly decreased PC(14:0/16:0) and PC(16:0/16:0) was observed in cisplatin-treated HEK293 and Cake-1 cells. Bromophenol lactone treatment prior to cisplatin exposure further decreased PC(14:0/16:0), plasmenylcholine(16:0/16:1), and plasmenylcholine(16:0/18:1) in HEK293 and inhibited cisplatin-induced increases in plasmenylcholine(16:1/22:6) in Caki-1. Treatment with bromophenol lactone prior to cisplatin exposure also increased several arachidonic containing phospholipids including PC(16:0/20:4), PC(18:1/20:4), and PC(18:0/20:4) versus cisplatin only treatment. These results demonstrated that phospholipase A2 inhibition protected against chemotherapy-induced cell death in multiple human renal cell lines and also identified phospholipids that were specifically altered during cell death. The results further demonstrated that alterations in these phospholipids correlated with protection against cell death in the presence of phospholipase A2 inhibitors. Masood and coworkers used normal and reversed-phase LC-MS/MS to quantitate multiple sphingolipid classes in HEK293 cells [151]. These results showed that more than 75% of the ceramides, monohexo sylceramides, and SM exist as d18:1Δ4 c16:0, d18:1Δ4 c24:1, and d18:1-4 c24:0.
5. CONCLUDING REMARKS AND PERSPECTIVES
Novel lipidomics is an emerging methodology that holds promise for a systematic and comprehensive study of lipids and their derivatives in health and disease. Various kidney diseases are associated with significant changes in metabolism and plasma concentration of lipids and lipoproteins, as well as lipid-related metabolites and metabolic pathways. These changes play an important role in the pathogenesis of local and systemic inflammation, impaired energy metabolism, and progression of kidney disease. The combination of lipid profiling and multivariate statistics is useful for the discovery of potential biomarkers and novel therapeutic modalities, as well as monitoring response to therapeutic intervention.
Recent advances in MS-based technologies and rapid improvements in chromatography, especially UPLC–MS combined with bioinformatics, have improved our understanding of the role of lipid-derived metabolites in the pathogenesis and progression of kidney disease. Although currently available tools allow identification of lipid-derived metabolite structure with high resolution, further advances in analytical techniques and data handling are clearly needed for more effective data preprocessing, data mining, statistical analysis, biomarker identification, and interpretation of biochemical pathways.

cistanche extract: better kidney function
ACKNOWLEDGMENTS
This study was supported by the Program for New Century Excellent Talents in University (NCET-13-0954) and Changjiang Scholars and Innovative Research Team in University (IRT1174) from the Ministry of Education of China, National Natural Science Foundation of China (J1210063, 81202909, 81274025, 81001622), the project “As a Major New Drug to Create a Major National Science and Technology Special” (2014ZX09304307- 002), China Postdoctoral Science Foundation (2012M521831, 2014T70984), National Innovation Training Plan Program (201310697004), Key Program for the International S&T Cooperation Projects of Shaanxi Province (2013KW31-01), Natural Science Foundation of Shaanxi Provincial Education Department (2013JK0811), and the Administration of Traditional Chinese Medicine of Shaanxi (13-ZY006).
*Key Laboratory of Resource Biology and Biotechnology in Western China, Ministry of Education, The College of Life Sciences, Northwest University, Xi’an, Shaanxi, PR China
†Division of Nephrology and Hypertension, School of Medicine, University of California, Irvine, California, USA
{School of Chinese Materia Medica, Beijing University of Chinese Medicine, Beijing, PR China
REFERENCES
[1]A. Levin, N.R. Powe, J. Rosset, et al., Chronic kidney disease as a global public health problem: approaches and initiatives—a position statement from Kidney Disease Improving Global Outcomes, Kidney Int. 72 (2007) 247–259.
[2]K. Makris, N. Kafkas, Neutrophil gelatinase-associated lipocalin in acute kidney injury, Adv. Clin. Chem. 58 (2012) 141–191.
[3]X.B. Ling, E.D. Mellins, K.G. Sylvester, H.J. Cohen, Urine peptidomics for clinical biomarker discovery, Adv. Clin. Chem. 51 (2010) 181–213.
[4]T.K. Sigdel, R.B. Klassen, M.M. Sarwal, Interpreting the proteome and peptidome in transplantation, Adv. Clin. Chem. 47 (2009) 139–169.
[5]N.D. Vaziri, Dyslipidemia of chronic renal failure: the nature, mechanisms and potential consequences, Am. J. Physiol. Renal Physiol. 290 (2006) 262–272.
[6]N.D. Vaziri, J. Yuan, Z. Ni, S.B. Nicholas, K.C. Norris, Lipoprotein lipase deficiency in chronic kidney disease is accompanied by down-regulation of endothelial GPIHBP1 expression, Clin. Exp. Nephrol. 16 (2012) 238–243.
[7]N.D. Vaziri, Molecular mechanisms of lipid disorders in nephrotic syndrome, Kidney Int. 63 (2003) 1964–1976.
[8]N.D. Vaziri, Lipotoxicity and impaired HDL-mediated reverse cholesterol/lipid transport in chronic kidney disease, J. Ren. Nutr. 20 (2010) S35–S43.
[9]N.D. Vaziri, K. Norris, Lipid disorders and their relevance to outcomes in chronic kidney disease, Blood Purif. 31 (2011) 189–196.
[10]N.D. Vaziri, Role of dyslipidemia in impairment of energy metabolism, oxidative stress, inflammation and cardiovascular disease in chronic kidney disease, Clin. Exp. Nephrol. 18 (2014) 265–268.
[11]C. Waldner, G. Heise, K. Schroer, P. Heering, COX-2 inhibition and prostaglandin receptors in experimental nephritis, Eur. J. Clin. Invest. 33 (2003) 969–975.
[12]A. Hartner, A. Pahl, K. Brune, M. Goppelt-Strube, Upregulation of cyclooxygenase-1 and the PGE2 receptor EP2 in rat and human mesangial proliferative glomerulonephritis, Inflamm. Res. 49 (2000) 345–354.
[13]S. Tomasoni, M. Noris, S. Zappella, et al., Upregulation of renal and systemic cyclooxygenase-2 in patients with active lupus nephritis, J. Am. Soc. Nephrol. 9 (1998) 1202–1212.
[14]C. Zoja, A. Benigni, M. Noris, et al., Mycophenolate mofetil combined with a cyclooxygenase-2 inhibitor ameliorates murine lupus nephritis, Kidney Int. 60 (2001) 653–663.
[15]T. Takano, A.V. Cybulsky, W.A. Cupples, et al., Inhibition of cyclooxygenases reduces complement-induced glomerular epithelial cell injury and proteinuria in passive Heymann nephritis, J. Pharmacol. Exp. Ther. 305 (2003) 240–249.
[16]G. Heise, B. Grabensee, K. Schro€r, P. Heering, Different actions of the cyclooxygenase 2 selective inhibitor flosulide in rats with passive Heymann nephritis, Nephron 80 (1998) 220–226.
[17]Z.G. Xu, S.L. Li, L. Lanting, et al., Relationship between 12/15-lipoxygenase and COX-2 in mesangial cells: potential role in diabetic nephropathy, Kidney Int. 69 (2006) 512–519.
[18]A. Dey, R.S. Williams, D.M. Pollock, et al., Altered kidney CYP2C, and cyclooxygenase-2 levels are associated with obesity-related albuminuria, Obes. Res. 12 (2004) 1278–1289.
[19]X. Zhao, J.E. Quigley, J. Yuan, et al., PPAR-alpha activator fenofibrate increases renal CYP-derived eicosanoid synthesis and improves endothelial dilator function in obese Zucker rats, Am. J. Physiol. 290 (2006) H2187–H2195.
[20]Y. Zhou, S. Lin, H.H. Chang, et al., Gender differences of renal CYP-derived eicosanoid synthesis in rats fed a high-fat diet, Am. J. Hypertens. 18 (2005) 530–537.




