Quercetin Improves Myocardial Redox Status in Rats With Type 2 Diabetes
Mar 26, 2022
For more information, contact tina.xiang@wecistanche.com
Objective. Emerging data indicate that oxidative stress is closely associated with the pathogenesis of cardiovascular disease in type 2 diabetes mellitus(T2DM). The present study aimed to assess the effect of the most abundant flavonoid in the human diet quercetin (Q) on the myocardial redox status in rats with T2DM.
Methods. T2DM was induced in male Wistar rats by a high caloric diet (for 14 weeks) and two streptozotocin (25 mg/kg b.w.) injections applied in four weeks of the diet, once a week for two weeks. The Q was administered intragastrically by gavage in a dose of 10 or 50 mg/kg of the bodyweight for 8 weeks starting from the 8th day after the last streptozotocin injection. The control rats received citrate buffer and seven days after the last STZ injection, basal glucose levels were measured in all animals.
Results. Administration of Q increased insulin sensitivity in diabetic rats with a more pronounced effect at a dose of50 mg/kg b.w. The Q also decreased free radical oxidation in the heart mitochondria of diabetic animals, thus limiting the formation of advanced oxidation protein products in a dose-dependent manner and normalized the activity of antioxidant enzymes(superoxide dismutase, glutathione peroxidase, glutathione reductase) in cardiac mitochondria independently of the dose used. In addition, the Q in both doses prevented the development of oxidative stress in the T2DM rat's cardiomyocytes by reducing NADPH oxidase and xanthine oxidase activities.
Conclusions. The findings demonstrate that Q in both doses 10 mg/kg and 50 mg/kg can protect from the development of oxidative stress in cardiomyocytes in diabetic rats. The present data indicate that the use of Q may contribute to the amelioration of cardiovascular risk in patients with T2DM.
Keywords: quercetin, type 2 diabetes mellitus, cardiomyocytes, oxidative stress

Diabetes mellitus (DM) is one of the fastest-growing global health emergencies of the21* century. According to the International Diabetes Federation, 463 million people worldwide had diabetes in 2019, and this number is assumed to reach 700 million by the year 2045.DM is associated with a wide range of cardiovascular conditions that collectively increase the risk of cardiovascular death by 132% in diabetics compared to individuals without DM(IDF Diabetes Atlas 2019).
The DM is known to cause specific changes in the myocardium structure and function independent of coronary artery disease or hypertension is commonly defined as diabetic cardiomyopathy (DCM).DCM is manifested by diastolic dysfunction followed by abnormalities in systolic function, which can lead to heart failure (Liu et al.2014). It is also known that metabolic disturbances in cardiomyocytes at DCM(hypertrophy, inflammation, apoptosis, fibrosis) are closely associated with the development of oxidative stress(Kayama et al.2015).
Reactive oxygen species (ROS), including superoxide anion, hydroxyl radical, and hydrogen peroxide are critical signaling molecules with important roles in both cardiac physiology and disease. Both cytosolic sources, including NADPH oxidases (NOX), xanthine oxidase(XO), cyclooxygenases, and cytochrome P450 enzymes, and mitochondrial sources, such as the respiratory chain and monoamine oxidases, contribute to the intracellular ROS pool. Under physiological conditions, cardiac ROS signaling regulates the heart development and cardio-myocyte maturation, cardiac calcium handling, excitation-contraction coupling, and vascular tone. However, pathological conditions of unregulated ROS production lead to elevated ROS levels, which can result in oxidative stress through oxidative damage of DNA, proteins, and lipids, as well as mitochondrial dysfunction and cell death (Peoples et al. 2019). Indeed, dysregulated ROS production and oxidative stress have been implicated in a lot of cardiac diseases, including diabetic cardiomyopathy (Ritchie and Abel 2020).
The excessive ROS production by mitochondria can directly affect the contractile function of the myocardium by oxidative modification of enzymes and ion channels that participate in the regulation of the excitation-relaxation cycle (Brown and Griendling 2015; Bai et al. 2016).In addition, ROS stimulates the proliferation of cardiomyocytes and activates fibroblasts and matrix metalloproteinases, which lead to hypertrophy and remodeling of the myocardium resulting in the formation of heart failure (DOria et al.2020). Therefore, it is suggested that pharmacological action aimed at reducing ROS over-production may be effective for the prevention and correction of diabetic cardiovascular complications.
At present, it has been established that dietary intake of natural polyphenols, particularly flavonoids, can reduce the risk of diabetes and cardiovascular diseases and the protective mechanisms may not only be dependent on the antioxidant and anti-inflammatory effects of flavonoids, but also include their properties affecting glycemia, glucose utilization, and insulin secretion(Ghorbaniet al.2019; Lutz et al.2019).
Quercetin (Q;3,5,7,34-pentahydroxy flavon)is the most abundant flavonoid in the human diet. There is a wide range of biological functions of Q including effects on pathways involved in inflammation, diabetes, and cardiovascular diseases(Zahedi et al. 2013). Several studies have demonstrated the mechanisms of the antidiabetic effects of the Q, including
decreased lipid peroxidation, increased activity of antioxidant enzymes (e.g., superoxide dismutase, SOD; glutathione peroxidase, GPX; catalase, CAT), reduced absorption of glucose in the intestine due to inactivation of the transporter GLUT2 and others (Shi et al.2019; Salehi et al.2020). The cardioprotective effect of Q on differentiated cardiomyocytes, due to the inactivation of mitogen-activated protein kinase and protein kinase B signaling pathways, has also been shown (Daubney et al.2015).
Future research devoted to understanding the cardioprotective action mechanisms in diabetes could contribute to diabetes care improvement.
The aim of the present work was to study the effect of quercetin on the redox status of cardiomyocytes in rats with type2 diabetes mellitus(T2DM).

Materials and methods
Chemicals. All chemicals used were of analytical reagent grade quality and purchased from Sigma Chemical Co.(St.Louis, MO, USA). Quercetin was provided by the PJSCSICBorshchahivskiy Chemical-Pharmaceutical Plant(Kyiv, Ukraine), and contained a minimum of 96.5% of the active substance.
Experimental design. The present study was approved by the bioethics committee of the"V. Dani-levsky Institute for Endocrine Pathology Problems of National Academy of Medical Sciences of Ukraine"(Kharkiv, Ukraine) and performed in accordance with the European Convention for the Protection of Vertebrate Animals Used for Experimental and Other Scientific Purposes(Strasbourg, 1986).
The experiments were performed on 32 male Wistar rats(12-week-old, 200-230 g body weight, b.w.), which were housed in Plexiglas cages(3 animals per cage) at a temperature of 22±1℃C in a constant 12-hour light/dark cycle.
The animal model of T2DM induced by a high caloric diet combined with multiple low dose streptozotocin (STZ) injections were used. The model provides the development of two main features of T2DM: the high caloric diet initiates insulin resistance and the low-dose STZ induces a mild impairment of insulin secretion. Thus, the model mimics the natural history of the disease events(from insulin resistance to β-cell dysfunction) as well as the metabolic characteristics of human T2DM(Skovso 2014).
Control intact rats (n=8) were fed a standard diet for 14 weeks. Experimental rats(n=24) were fed the high-calorie diet containing 15% lard, 25% sucrose, 1% bile salts, and 59% standard feed for 14 weeks. The animals had free access to water. In four weeks, experimental rats were intraperitoneally(.p.injected with STZ(25 mg/kg b.w.)once per week for two weeks(Lin et al. 2010). Control rats received citrate buffer following the same scheme. Seven days after the last STZ injection, basal glucose was measured in all animals, and experimental rats were divided into three groups: untreated diabetic rats (Diabetes, n=8) and diabetic rats treated with Qin a dose of 10 mg/kg b.w.(Diabetes+Q10,n=8)or 50 mg/kg b.w. (Diabetes+Q50, n=8)once per day intragastrically by gavage for8 weeks after diabetes induction. Untreated diabetic rats received vehicles along with the same scheme.
The animals were sacrificed according to the protocol of the ethics committee.
Measurement of glucose homeostasis. The intraperitoneal insulin tolerance test (IPITT) was performed in overnight fasted rats. Blood glucose concentrations were initially measured at the basal condition(0 min), then the animals were given ani. p. injection of insulin 0.25 U/kg b.w.(Actrapid, Novo Nordisk, Agsvaerd, Denmark)followed by an i.p. injection of glucose (2 g/kg)(Bowe et al.2014). Subsequently, tail blood glucose levels were measured using a glucose analyzer Eksan-G(Analita Firm Joint Stock Company Ltd., Vilnius, Republic of Lithuania) at 15, 30,60,90, and 120 min after the glucose load.
Isolation of mitochondria. Mitochondria were isolated by conventional procedures. Freshly excised rat heart was homogenized in a medium containing 0.18 M KCl, 10 mM EDTA, 0.5% bovine serum albumin(BSA),10mM HEPES,pH7.4.Homogenates were cleared from debris and nuclei by two times centrifugation at 500 g(10 min at 4°C). Mitochondria were pelleted from the supernatants at 10000g (15 min at 4°C) and resuspended in an isolation buffer.
The post mitochondrial supernatant fractions were used for the determination of XO and NOX activities. To remove EDTA and albumin, mitochondrial pellets were centrifuged at 10000g(15 min,4°C) and resuspended in wash buffer containing 0.18 M KCl and 10mM HEPES(pH7.4) twice (DiLisa et al.1994).
Spectrophotometry. All spectrophotometry was performed using double-beam UV-VIS spectrophotometer Shimadzu UV-1800 (Shimadzu Corporation, Kyoto, Japan).
Measurement of advanced oxidation protein products (AOPP). Determination of AOPP levels was performed by the modified method of Witko-Sarsat and colleagues (Sagor et al.2015). The heart mitochondria(0.2 mg protein) were treated with0.1%Triton-X100 and diluted in PBS(pH7.4)solution.KI (1.16 M) was added to the assay medium, followed 2 min later by the addition of glacial acetic acid. The absorbance of the reaction mixture was immediately read at 340 nm against the blank. AOPP concentrations were expressed as nmol-equivalent chloramine T/mg protein.
Measurement of enzyme activity. NOX activity was examined as NADPH-dependent O2 production by post mitochondrial supernatant fraction using SOD-inhibitable ferricytochrome c reduction (Li et al. 2002). The postmitochondrial fraction of the heart homogenate(1 mg protein) was supplemented with 10mM Tris-HCl(pH7.8),500μMcytochrome c,100μM NADPH, was incubated at 37℃C for 30 min with or without 200 U/ml SOD. Cytochrome reduction was measured by reading absorbance at 550 nm (8=21 mM-1 cm-).
XO activity was assayed in air-equilibrated PBS (pH7.4)solution that contained a postmitochondrial fraction of the heart homogenate (1 mg protein)at 37°C after addition of xanthine(final concentration 360μM)by measurement of uric acid production from the change in absorbance at 295nm (e=9500M-'cm-') (Lee et al.2014).
Mn-SOD activity in the mitochondrial suspension was detected by inhibition of nitroblue tetrazolium (NBT)reduction caused by xanthine-XO system as O2 generator and the absorbance was finally deter-mined at 560 nm. The reaction mixture contained heart mitochondria (0.02 mg protein),0.05M carbonate buffer(pH10.2),0.1 mM EDTA,0.1 mM xanthine,25μM NBT,1 mU/ml XO. The results were expressed in arbitrary units (a.u.)of SOD activity per mg protein. One a.u.means the enzyme amount causing 50%inhibition of NBT reduction rate (Wang et al. 2018).
Glutathione reductase (GR) activity was measured by tracking the oxidation of NADPH at 340 nm in the reaction of oxidized glutathione (GSSG) reduction. To perform the reaction, mitochondria from the heart (0.1 mg protein) were incubated for 10 min at 37°℃ in assay buffer containing 20 mM Tris-HCl (pH7.4),0.25 mM EDTA,10 μM FAD,3 mM GSSG and 0.1 mM NADPH(Raza and John 2004).
The estimation of GPX activity was made based on the enzyme's ability to catalyze tertbutyl hydroperoxide degradation using reduced glutathione(GSH)as a substrate. Reaction mixture(pH7.0) contained heart mitochondria (0.1 mg protein), 0.56mM NADPH,1.0 U/ml GR,7.5 mM sodium azide,5 mM GSH,5 mM EDTA and 0.05 M phosphate buffer. Tert-butyl hydroperoxide (23 mM) was added to initiate the reaction. The rate of GSSG formation was determined by GR reaction from a decrease in the absorbance of probes at 340 nm (Antunes et al.2002).
Measurement of protein content in the samples. Mitochondrial protein was determined by the Lowry method modified by Miller, with BSA as the standard (Lowry et al.1951; Miller 1959).
Statistical analysis. Data are presented as mean±standard error of the mean (SEM). The Shapiro-Wilk test was used to test the normality of data distribution. For multiple comparisons of data with a normal distribution, a parametric one-way analysis of variance (ANOVA)was performed and the Student-Newman-Keuls method was used to test differences in means. Values were considered statistically significant at p<0.05.

Results
We revealed that the level of basal glucose in rats fed with a high caloric diet and injected with STZ was significantly higher than in the control group (Table 1). In addition, the areas under the glycemic curves (AUC) obtained during IPITT were almost three times larger in diabetic animals compared to control ones(Table 1, Figure 1). These data confirm the development of relative insulin deficiency and insulin resistance in experimental rats.
Administration of Q had no significant impact on the basal hyperglycemia of diabetic animals. However, Q affected insulin sensitivity in rats with T2DM in a dose-dependent manner, decreasing IPITT AUC by 27%in experimental group receiving 10 mg/kg of the drug, and by 41% in the group receiving 50 mg/kg of Q compared to animals receiving vehicle(Table 1, Figure 1).

 in control and diabetic rats. Data are shown as mean ± SEM (n=8); *р≤0.05 – statistical difference vs. Control; #р≤0.05 – statistical difference vs. Diabetes; †р≤0.05 – statistical difference vs. Diabetes+Q10")
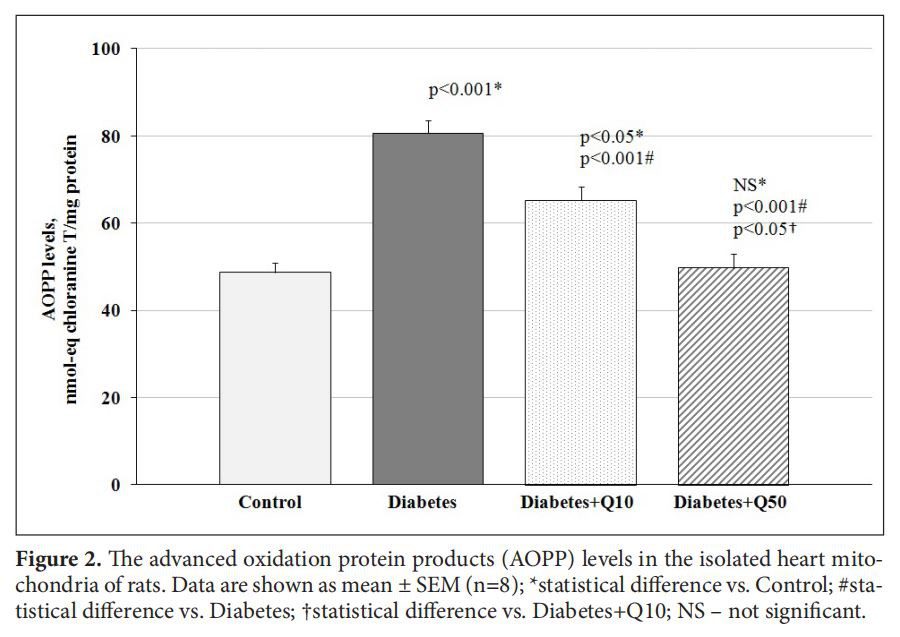
AOPP level in the heart mitochondria of experimental rats was increased by 66% confirming the development of oxidative stress in T2DM(Figure 2). Administration resulted in lowering of AOPP accumulation in the heart of diabetic animals depending on the dose of the drug. In the experimental group receiving 50 mg/kg of Q, the concentration of AOPP decreased to the level of the control group, suggesting antioxidant properties of Q(Figure 2).
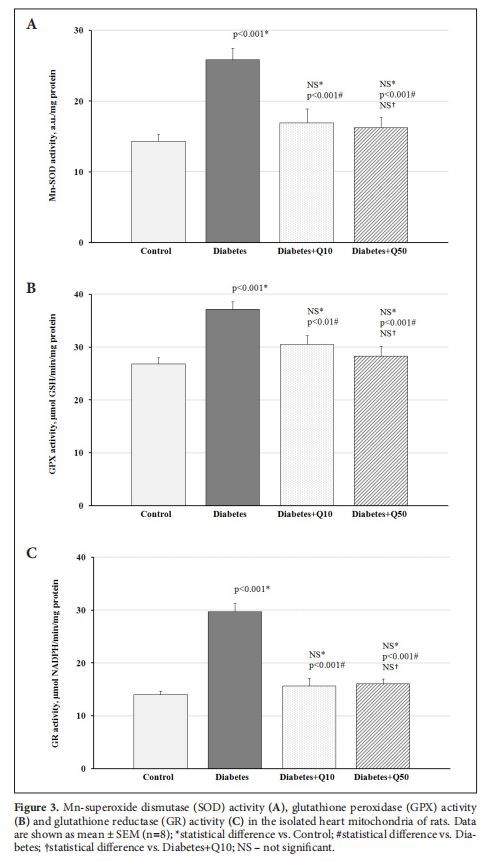
As can be seen in Figure 3, oxidative stress in the heart mitochondria of rats with T2DM was accompanied by the induced activity of antioxidant enzymes -Mn-SOD, GPO, and GR. The Q administration in both doses normalized the activities of studied anti-oxidant enzymes in diabetic animals to the level of the control group (Figure 3).
It was found that T2DM was associated with a 65%and 52% increase of NOX and XO activities, two primary cytosolic sources of O2 in cardiomyocytes, respectively, in the postmitochondrial fraction of the heart homogenate of experimental rats (Figure 4). The use of Q led to the normalization of NOX and XO activities in the heart of diabetic animals independently of dose (Figure 4).
 activity (A) and xanthine oxidase (XO) activity (B) in the postmitochondrial fraction of the heart homogenate of rats. Data are shown as mean ± SEM (n=8); *statistical difference vs. Control; #statistical difference vs. Diabetes; †statistical difference vs. Diabetes+Q10; NS – not significant.")
Discussion
At present, oxidative stress defined as a sustained increase in free radicals due to an imbalance in their production and utilization is thought to be a universal mechanism that combines the various pathogenetic pathways of diabetic cardiovascular complications(Huynh et al.2014).
Oxidative stress induced by hyperglycemia plays a leading role in the pathogenesis of T2DM(Shah and Brownlee 2016). The main pathological features of T2DM is reduced insulin secretion and decreased sensitivity to the hormone in peripheral tissues. At the first stage of our study, insulin resistance was modeled in rats using a high-calorie diet. During four weeks, rats receiving a high-calorie diet were. p. injected with STZtoreproduce a relative insulin deficiency. Elevated levels of basal glycemia and increased AUC in IPITT indicated the development of insulin deficiency and insulin resistance in model animals.
Administration of Q didn't affect the levels of basal hyperglycemia of diabetic rats, but improved insulin sensitivity in a dose-dependent manner. The results are consistent with data from other studies on the ability of Q to reduce insulin resistance(Shi et al. 2019). The Q increases glucose utilization in peripheral tissues and reduces hepatic glucose production. The flavonoid is reported to activate an insulin-independent mechanism involving AMP-dependent protein kinase, which stimulates the glucose transporter GLUT4 translocation into the plasma membrane in skeletal muscle and downregulates the key gluconeogenesis enzymes in the liver(phosphoenol-pyruvate carboxykinase and glucose 6-phosphatase)(Eid and Haddad 2017). The improvement of insulin sensitivity by Q may also be due to its ability to increase adiponectin expression in the white adipose tissue independently of the expression of peroxisome proliferator-activated receptors α(Wein et al. 2010).
The antidiabetic effect of Q can also be realized by reducing the absorption of glucose in the intestine through the inactivation of the glucose transporter GLUT2 (Gaueret al.2018)and inhibition of the intestinal α-glucosidases activity(Pereira et al. 2011). In addition, the Q has been shown to inhibit the enzyme 1lβ-hydroxysteroid dehydrogenase-1, which plays a key role in the development of obesity and T2DM by affecting the action of glucocorticoid hormones in the liver, adipose tissue, and pancreatic β-cells (Torres-Piedra et al.2010).
It is assumed that both the development and progression of diabetic complications are due to an increase in the production of ROS(Kayama et al. 2015).In cardiomyocytes, ROSare generated by mitochondrial electron transport chain (ETC), NOX, and XO(Tsutsui et al.2011). It can be argued that mitochondria are the main oxidative stress contributors to T2DM(Shah and Brownlee 2016). The develop-ment of oxidative stress in mitochondria is accompanied by disruption of redox-sensitive signaling pathways, decreased oxidative phosphorylation and ATP production, damage of mitochondrial DNA, and, by consequence, a further increase in ROS production. As a result, the pathogenic cascade of molecular events, which amplifies itself, leads to the launch of proapoptotic signaling pathways and cell death. Given the importance of mitochondria in cell metabolism, their dysfunction may lead to various diseases including cardiovascular pathology. Mitochondrial dysfunction occurs in patients with coronary heart disease, cardiomyopathy, heart failure, stroke, and T2DM (Bai et al.2016).
In our study, the development of oxidative stress in the heart mitochondria of diabetic rats is evidenced by the accumulation of AOPP. The concentration of AOPP is a relatively novel marker of oxidative stress, which reflects the overall redox status of proteins in cells and tissues. AOPP are reported to be elevated in patients with DM, cardiovascular diseases, hypertension, atherosclerosis(Conti et al.2019), and the level correlates with insulin resistance and the presence of severe diabetic complications(Gradinaru et al. 2013; Taylor et al.2015). Oxidative protein modifications play a key role in the development of diabetic myocardial dysfunction. Redox changes of protein structures may result in dissociation of catalytic subunits of enzymes, unfolding, aggregation, or fragmentation of proteins involved in contractility, excitation-contraction coupling, protein folding, antioxidant defense, metabolism, and Ca2* handling in the diabetic heart(Varga et al. 2015). It has also been found that AOPP induces proapoptotic signaling in cardiomyocytes leading to the development of DCM (Zhang et al.2016).
In animals treated with Q in a dose of 10 mg/kg, a significant reduction of AOPP in the heart mitochondria was observed, whereas a dose of 50 mg/kg of Q completely normalized this marker as compared to rats that received vehicle.
The antioxidant effect of the Q in mitochondria may include several main mechanisms. Flavonoids have the ability to scavenge ROS due to the hydroxyl configuration of the B-ring in the structure(Kumar and Pandey 2013). The direct scavenging of O,-generated in mitochondrial complex II of the ETC in isolated rat heart mitochondria has been shown for Q(Dudy-lina et al. 2019). Besides, flavonoids can attenuate the formation of ROS in mitochondria through the inhibition of complex I and III of the ETC. Q inhibition of complex I activity significantly decreased H2O2, production in isolated rat heart mitochondria (Lagoa et al.2011). In addition, the reduction in mitochondrial ROS generation by flavonoids was found to be associated with partial uncoupling properties, particularly, in cardiac mitochondria (Bernatoniene et al.2014). Furthermore, flavonoids have been shown to stimulate mitochondrial biogenesis through the induction of transcriptional co-activator PGC-la and its upstream regulator SIRT1. The latter leads to stimulation of a series of transcription factors(NRF1, NRF2, and estrogen-related-a), involved in the regulation of energy metabolism and preservation of homeostasis in mitochondria(Kicinska and Jarmuszkiewicz 2020).

As a result of our experiment, the increase of oxidative stress in the heart mitochondria of diabetic rats was accompanied by induction of the activity of antioxidant enzymes-Mn-SOD, GPO, and GR. It is well established, that a major mechanism in the cellular defense against oxidative stress is the activation of the Keapl/Nrf-2-dependent signaling pathway, which controls the expression of genes whose protein products are involved in the detoxication and elimination of reactive oxidants (Nguyen et al. 2009). In this regard, the observed induction of antioxidant enzymes in the heart of experimental animals is the implementation of negative feedback aimed to maintain redox homeostasis.
Administration of Q in both doses led to a decrease in the activity of Mn-SOD, GPO, and GR in the cardiac mitochondria of experimental animals to values statistically indifferent from the control group. Though Q is known to induce the activity of the above enzymes (Shi et al.2019), our results indicate that the antioxidant effect of Q in cardiomyocytes in T2DM is realized more by its ability to prevent hyperproduction of ROS and by consequence the development of mitochondrial dysfunction, than by stimulation of antioxidant defense system. Previously, it was shown that Q use increases mitochondria oxidative capacity and the activity of some enzymes of the Krebs cycle in the heart of rats with T2DM(Gorbenko et al.2019).
As noted above, mitochondria are the primary, but not the only source of ROS in cardiomyocytes. The second important factor in the development of oxidative stress in the diabetic heart is NOX, with NOX2 and NOX4 as the major myocardial isoforms. The main function of NOX is to generate ROS. NOX2 is localized in the plasma membrane and participates in intracellular redox signaling of the angiotensin II receptor and others, while NOX4 particularly has a mitochondrial localization signal and exhibits constitutive activity (Cave et al. 2005). It has been demonstrated that incubation of rat cardiomyocytes with early-glycated proteins stimulated ROS production through a protein kinase C(PKC)-dependent activation of NOX2(Zhang et al. 2006). Furthermore, cardiac hypertrophy, fibrosis, and apoptosis were found to be associated with upregulated NOX2 in the heart of mice with type 1 and type 2 DM (Ritchie et al.2007; Huynh et al.2013). An additional source of ROS in cardiomyocytes is XO, which catalyzes the oxidation of hypoxanthine and xanthine to uric acid using O, as an electron receptor and producing O, and H2O2, in the process. Although the contribution of XO reaction to the development of oxidative stress in diabetes has not been studied enough, XO inhibitors were shown to ameliorate cardiovascular dysfunction in the model of pacing-induced heart failure (Amado et al. 2005). In our study, T2DM led to a significant increase in NOX and XO activities in the heart of experimental rats. At the same time, Q administration independently from the dose-normalized the activity of enzymes. To date, several studies have shown the ability of Q to inhibit NOX activity by both reducing expressions of its subunits and preventing its activation through extracellular stimuli. The second mechanism could be realized by reducing the phosphorylation of the regulatory subunit p47phox due to the inhibitory effect of flavonoids on PKC (Redondo et al.2012; Jimenez et al. 2015). Besides, O and its metabolites are known to inhibit the activity of XO in a selective way, forming the conjugated three-ring structure with the active site and specific hydrogen-bonding interactions with catalytically relevant residues of the enzyme(Taka-Hama et al.201l; Cao et al.2014).
In summary, our study showed that Q in a dose-dependent manner decreased free radical oxidation in the myocardial mitochondria of rats with T2DM, thus limiting AOPP formation. In addition, the Q in a dose of both 10 mg/kg and 50 mg/kg prevented the development of oxidative stress in the heart of diabetic rats reducing NOX and XO activities. The obtained data suggest that the use of Q may contribute to the amelioration of cardiovascular risk in T2DM.





